

Fatty acid-binding protein 4, a point of convergence for angiogenic and metabolic signaling pathways in endothelial cells
- PMID: 24939870
- PMCID: PMC4132814
- DOI: 10.1074/jbc.M114.576512
Fatty acid-binding protein 4, a point of convergence for angiogenic and metabolic signaling pathways in endothelial cells
Abstract
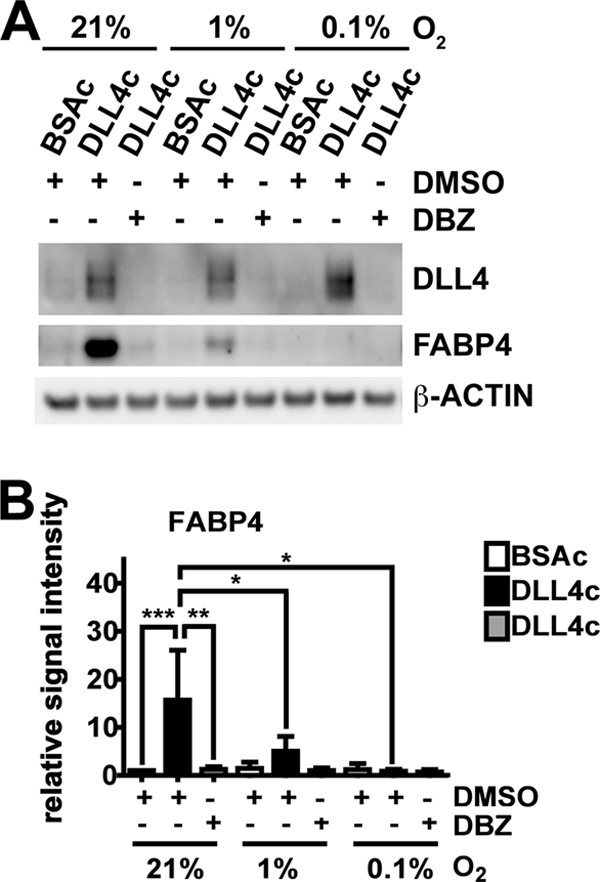
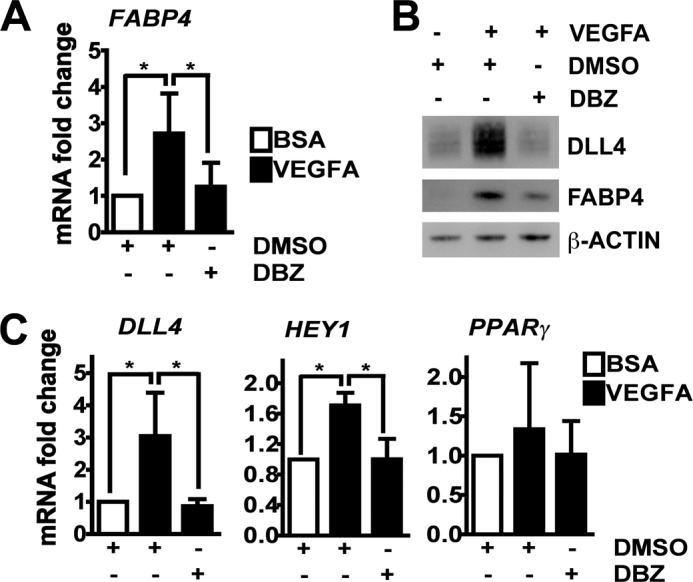
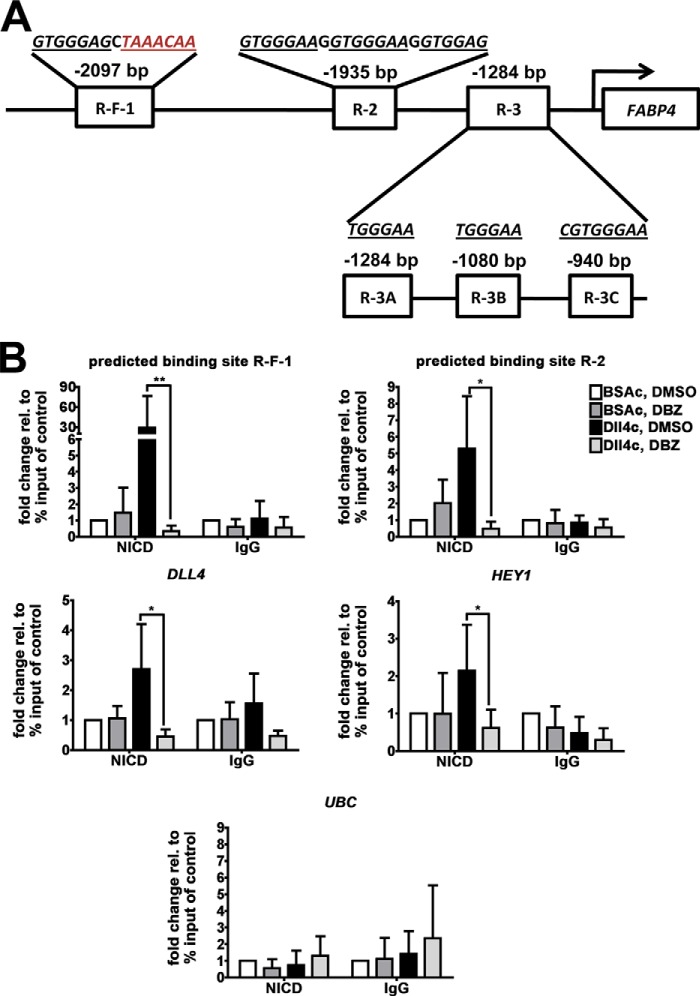
Fatty acid-binding protein 4 (FABP4) is an adipogenic protein and is implicated in atherosclerosis, insulin resistance, and cancer. In endothelial cells, FABP4 is induced by VEGFA, and inhibition of FABP4 blocks most of the VEGFA effects. We investigated the DLL4-NOTCH-dependent regulation of FABP4 in human umbilical vein endothelial cells by gene/protein expression and interaction analyses following inhibitor treatment and RNA interference. We found that FABP4 is directly induced by NOTCH. Stimulation of NOTCH signaling with human recombinant DLL4 led to FABP4 induction, independently of VEGFA. FABP4 induction by VEGFA was reduced by blockade of DLL4 binding to NOTCH or inhibition of NOTCH signal transduction. Chromatin immunoprecipitation of the NOTCH intracellular domain showed increased binding to two specific regions in the FABP4 promoter. The induction of FABP4 gene expression was dependent on the transcription factor FOXO1, which was essential for basal expression of FABP4, and FABP4 up-regulation following stimulation of the VEGFA and/or the NOTCH pathway. Thus, we show that the DLL4-NOTCH pathway mediates endothelial FABP4 expression. This indicates that induction of the angiogenesis-restricting DLL4-NOTCH can have pro-angiogenic effects via this pathway. It also provides a link between DLL4-NOTCH and FOXO1-mediated regulation of endothelial gene transcription, and it shows that DLL4-NOTCH is a nodal point in the integration of pro-angiogenic and metabolic signaling in endothelial cells. This may be crucial for angiogenesis in the tumor environment.
Keywords: Angiogenesis; DLL4; Endothelial Cell; FOXO; Fatty Acid-binding Protein; Metabolism; Notch Pathway; Vascular Endothelial Growth Factor (VEGF).
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






References
-
- Kralisch S., Fasshauer M. (2013) Adipocyte fatty acid-binding protein: a novel adipokine involved in the pathogenesis of metabolic and vascular disease? Diabetologia 56, 10–21 - PubMed
-
- Jenkins-Kruchten A. E., Bennaars-Eiden A., Ross J. R., Shen W. J., Kraemer F. B., Bernlohr D. A. (2003) Fatty acid-binding protein-hormone-sensitive lipase interaction. Fatty acid dependence on binding. J. Biol. Chem. 278, 47636–47643 - PubMed
-
- Nieman K. M., Kenny H. A., Penicka C. V., Ladanyi A., Buell-Gutbrod R., Zillhardt M. R., Romero I. L., Carey M. S., Mills G. B., Hotamisligil G. S., Yamada S. D., Peter M. E., Gwin K., Lengyel E. (2011) Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat. Med. 17, 1498–1503 - PMC - PubMed
-
- Holm S., Ueland T., Dahl T. B., Michelsen A. E., Skjelland M., Russell D., Nymo S. H., Krohg-Sørensen K., Clausen O. P., Atar D., Januzzi J. L., Aukrust P., Jensen J. K., Halvorsen B. (2011) Fatty acid-binding protein 4 is associated with carotid atherosclerosis and outcome in patients with acute ischemic stroke. PLoS One 6, e28785. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
