

Amyloids of alpha-synuclein affect the structure and dynamics of supported lipid bilayers
- PMID: 24940776
- PMCID: PMC4070068
- DOI: 10.1016/j.bpj.2014.05.001
Amyloids of alpha-synuclein affect the structure and dynamics of supported lipid bilayers
Abstract
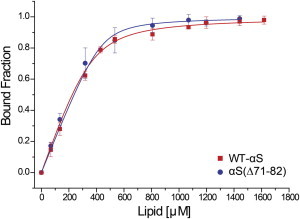
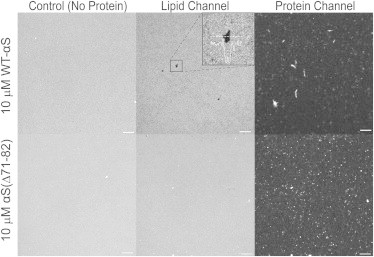
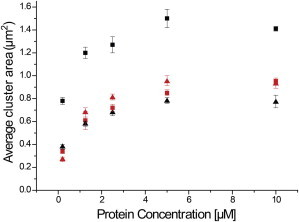
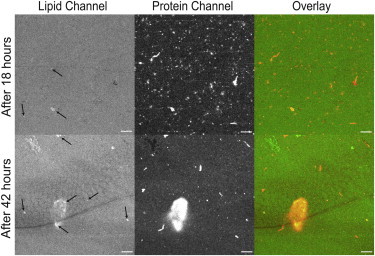
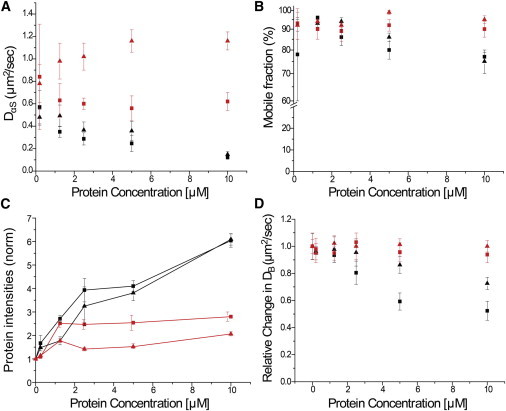
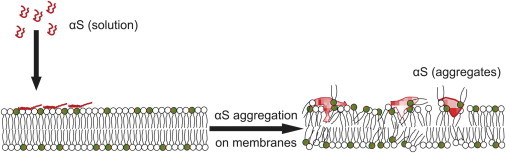
Interactions of monomeric alpha-synuclein (αS) with lipid membranes have been suggested to play an important role in initiating aggregation of αS. We have systematically analyzed the distribution and self-assembly of monomeric αS on supported lipid bilayers. We observe that at protein/lipid ratios higher than 1:10, αS forms micrometer-sized clusters, leading to observable membrane defects and decrease in lateral diffusion of both lipids and proteins. An αS deletion mutant lacking amino-acid residues 71-82 binds to membranes, but does not observably affect membrane integrity. Although this deletion mutant cannot form amyloid, significant amyloid formation is observed in the wild-type αS clusters. These results suggest that the process of amyloid formation, rather than binding of αS on membranes, is crucial in compromising membrane integrity.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- Spillantini M.G., Schmidt M.L., Goedert M. Alpha-synuclein in Lewy bodies. Nature. 1997;388:839–840. - PubMed
-
- Fiskum G., Starkov A., Chinopoulos C. Mitochondrial mechanisms of neural cell death and neuroprotective interventions in Parkinson’s disease. Ann. N. Y. Acad. Sci. 2003;991:111–119. - PubMed
-
- Polymeropoulos M.H., Lavedan C., Nussbaum R.L. Mutation in the α-synuclein gene identified in families with Parkinson’s disease. Science. 1997;276:2045–2047. - PubMed
-
- Farrer M.J. Genetics of Parkinson disease: paradigm shifts and future prospects. Nat. Rev. Genet. 2006;7:306–318. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
