

Lateral motion and bending of microtubules studied with a new single-filament tracking routine in living cells
- PMID: 24940780
- PMCID: PMC4070070
- DOI: 10.1016/j.bpj.2014.04.046
Lateral motion and bending of microtubules studied with a new single-filament tracking routine in living cells
Abstract
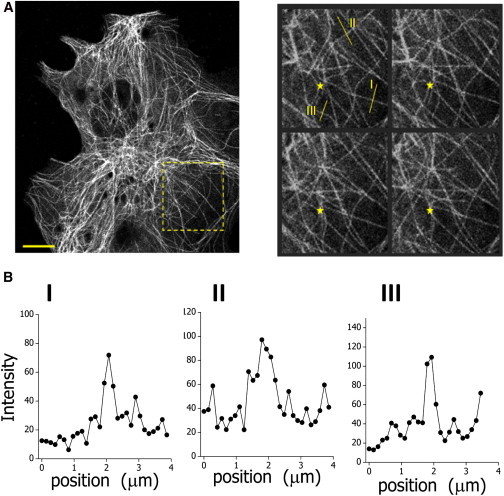
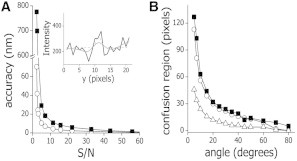
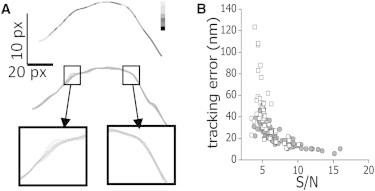
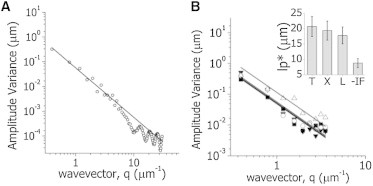
The cytoskeleton is involved in numerous cellular processes such as migration, division, and contraction and provides the tracks for transport driven by molecular motors. Therefore, it is very important to quantify the mechanical behavior of the cytoskeletal filaments to get a better insight into cell mechanics and organization. It has been demonstrated that relevant mechanical properties of microtubules can be extracted from the analysis of their motion and shape fluctuations. However, tracking individual filaments in living cells is extremely complex due, for example, to the high and heterogeneous background. We introduce a believed new tracking algorithm that allows recovering the coordinates of fluorescent microtubules with ∼9 nm precision in in vitro conditions. To illustrate potential applications of this algorithm, we studied the curvature distributions of fluorescent microtubules in living cells. By performing a Fourier analysis of the microtubule shapes, we found that the curvatures followed a thermal-like distribution as previously reported with an effective persistence length of ∼20 μm, a value significantly smaller than that measured in vitro. We also verified that the microtubule-associated protein XTP or the depolymerization of the actin network do not affect this value; however, the disruption of intermediate filaments decreased the persistence length. Also, we recovered trajectories of microtubule segments in actin or intermediate filament-depleted cells, and observed a significant increase of their motion with respect to untreated cells showing that these filaments contribute to the overall organization of the microtubule network. Moreover, the analysis of trajectories of microtubule segments in untreated cells showed that these filaments presented a slower but more directional motion in the cortex with respect to the perinuclear region, and suggests that the tracking routine would allow mapping the microtubule dynamical organization in cells.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Goldstein D., Elhanan T., Weihs D. Origin of active transport in breast-cancer cells. Soft Matter. 2013;9:7167–7173.
-
- Howard J. Sinauer Associates; Sunderland, MA: 2001. Mechanics of Motor Proteins and the Cytoskeleton.
-
- Mitchison T., Kirschner M. Dynamic instability of microtubule growth. Nature. 1984;312:237–242. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
