

DMXAA causes tumor site-specific vascular disruption in murine non-small cell lung cancer, and like the endogenous non-canonical cyclic dinucleotide STING agonist, 2'3'-cGAMP, induces M2 macrophage repolarization
- PMID: 24940883
- PMCID: PMC4062468
- DOI: 10.1371/journal.pone.0099988
DMXAA causes tumor site-specific vascular disruption in murine non-small cell lung cancer, and like the endogenous non-canonical cyclic dinucleotide STING agonist, 2'3'-cGAMP, induces M2 macrophage repolarization
Abstract
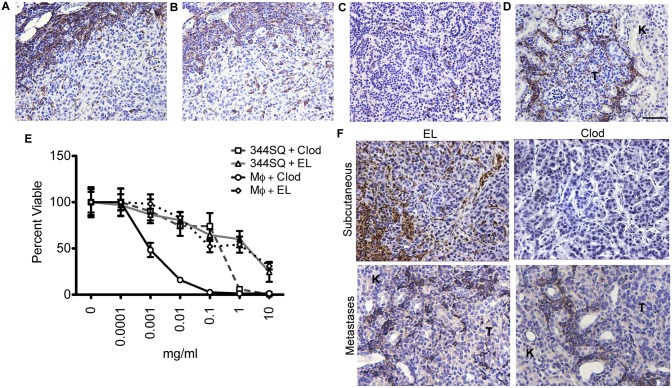
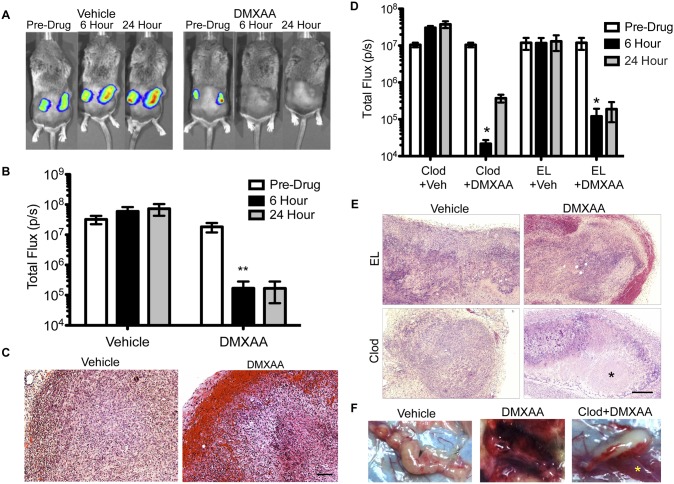
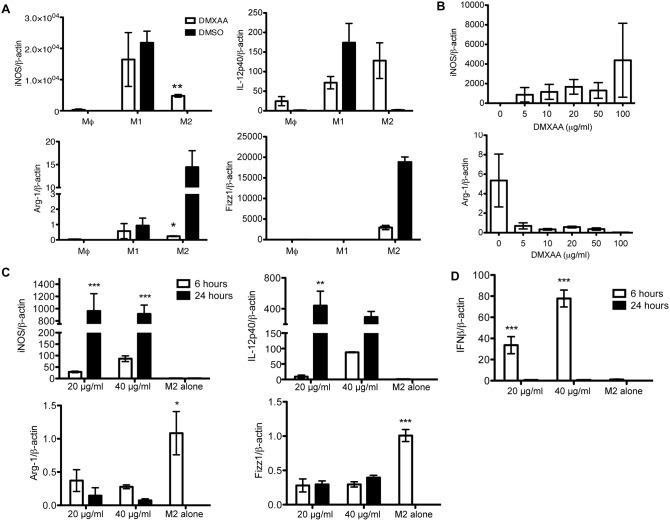
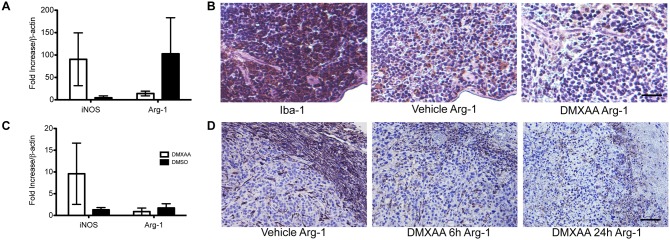
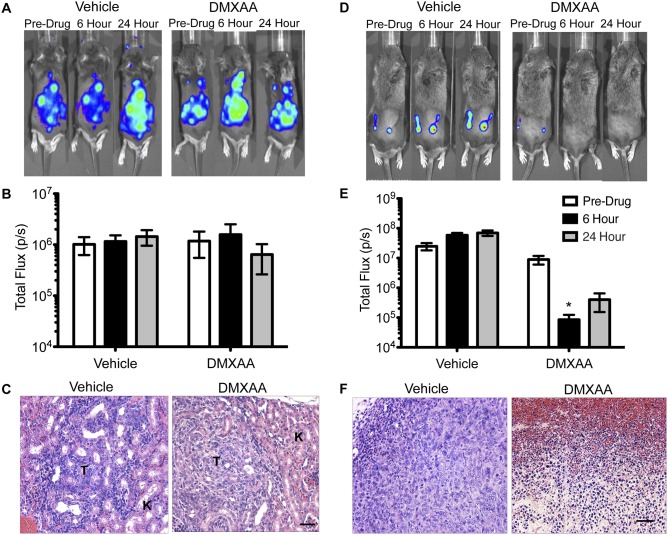
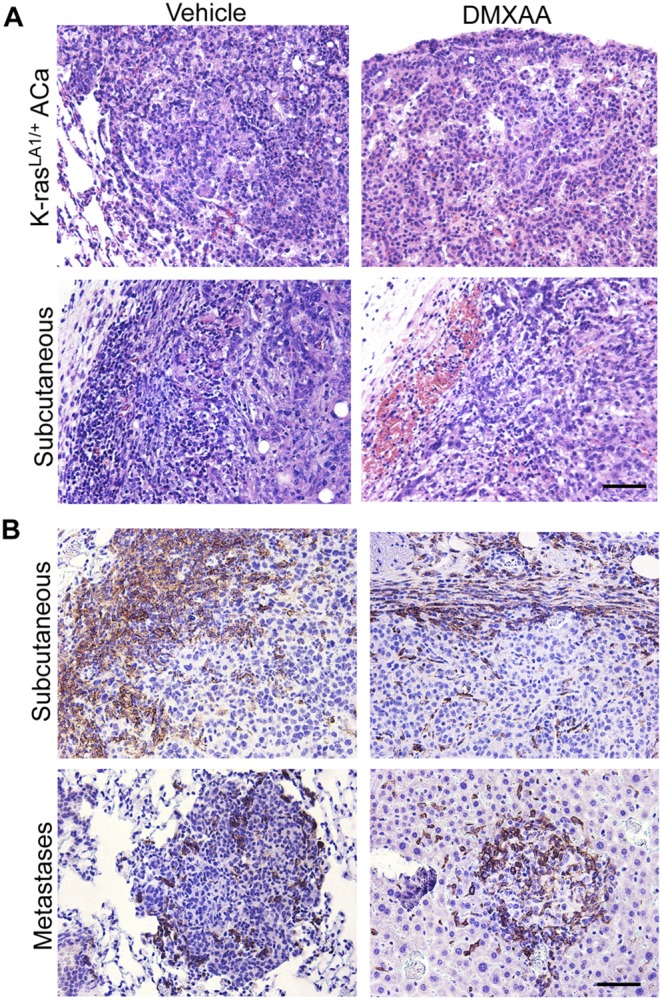
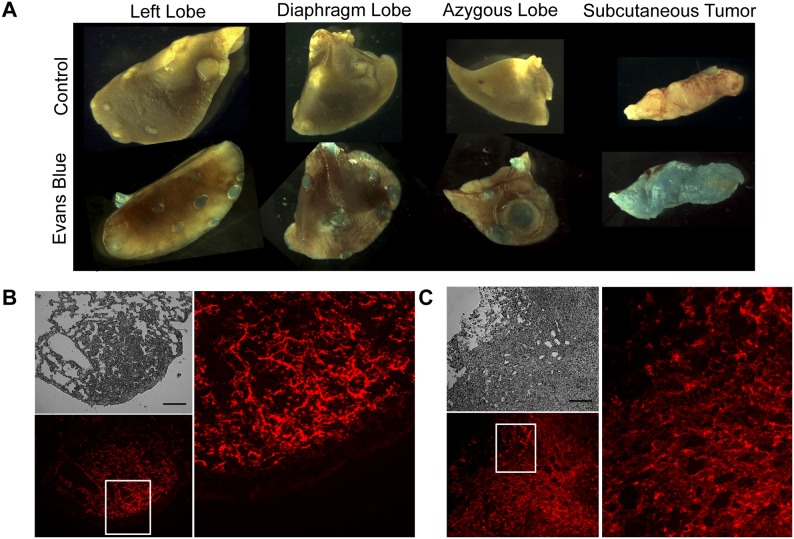
The vascular disrupting agent 5,6-dimethylxanthenone-4-acetic acid (DMXAA), a murine agonist of the stimulator of interferon genes (STING), appears to target the tumor vasculature primarily as a result of stimulating pro-inflammatory cytokine production from tumor-associated macrophages (TAMs). Since there were relatively few reports of DMXAA effects in genetically-engineered mutant mice (GEMM), and models of non-small cell lung cancer (NSCLC) in particular, we examined both the effectiveness and macrophage dependence of DMXAA in various NSCLC models. The DMXAA responses of primary adenocarcinomas in K-rasLA1/+ transgenic mice, as well as syngeneic subcutaneous and metastatic tumors, generated by a p53R172HΔg/+; K-rasLA1/+ NSCLC line (344SQ-ELuc), were assessed both by in vivo bioluminescence imaging as well as by histopathology. Macrophage-dependence of DMXAA effects was explored by clodronate liposome-mediated TAM depletion. Furthermore, a comparison of the vascular structure between subcutaneous tumors and metastases was carried out using micro-computed tomography (micro-CT). Interestingly, in contrast to the characteristic hemorrhagic necrosis produced by DMXAA in 344SQ-ELuc subcutaneous tumors, this agent failed to cause hemorrhagic necrosis of either 344SQ-ELuc-derived metastases or autochthonous K-rasLA1/+ NSCLCs. In addition, we found that clodronate liposome-mediated depletion of TAMs in 344SQ-ELuc subcutaneous tumors led to non-hemorrhagic necrosis due to tumor feeding-vessel occlusion. Since NSCLC were comprised exclusively of TAMs with anti-inflammatory M2-like phenotype, the ability of DMXAA to re-educate M2-polarized macrophages was examined. Using various macrophage phenotypic markers, we found that the STING agonists, DMXAA and the non-canonical endogenous cyclic dinucleotide, 2'3'-cGAMP, were both capable of re-educating M2 cells towards an M1 phenotype. Our findings demonstrate that the choice of preclinical model and the anatomical site of a tumor can determine the vascular disrupting effectiveness of DMXAA, and they also support the idea of STING agonists having therapeutic utility as TAM repolarizing agents.
Conflict of interest statement
Figures


References
-
- Ribatti D, Nico B, Crivellato E, Roccaro AM, Vacca A (2007) The history of the angiogenic switch concept. Leukemia 21: 44–52. - PubMed
-
- McKeage MJ, Baguley BC (2010) Disrupting established tumor blood vessels: an emerging therapeutic strategy for cancer. Cancer 116: 1859–1871. - PubMed
-
- Tozer GM, Kanthou C, Baguley BC (2005) Disrupting tumour blood vessels. Nat Rev Cancer 5: 423–435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
