

Differential coupling of KLF10 to Sin3-HDAC and PCAF regulates the inducibility of the FOXP3 gene
- PMID: 24944246
- PMCID: PMC4166759
- DOI: 10.1152/ajpregu.00085.2014
Differential coupling of KLF10 to Sin3-HDAC and PCAF regulates the inducibility of the FOXP3 gene
Abstract
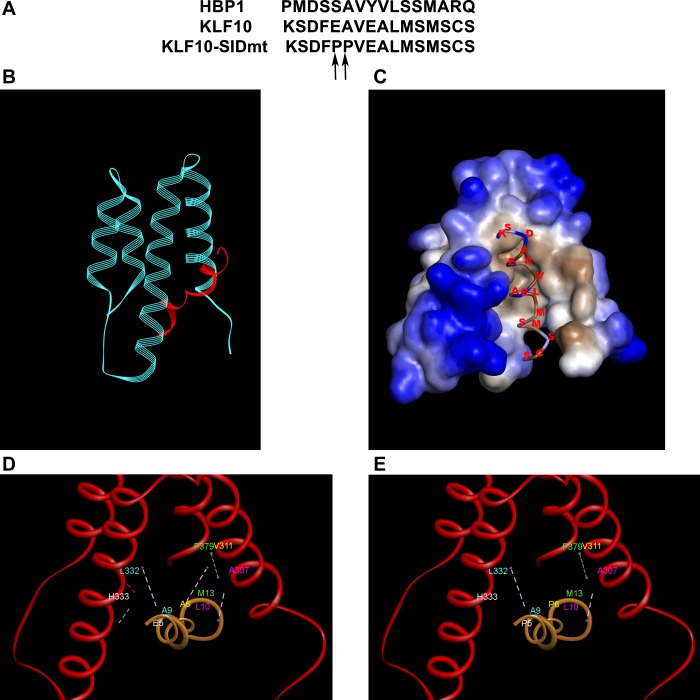
Inducible gene expression, which requires chromatin remodeling on gene promoters, underlies the epigenetically inherited differentiation program of most immune cells. However, chromatin-mediated mechanisms that underlie these events in T regulatory cells remain to be fully characterized. Here, we report that inducibility of FOXP3, a key transcription factor for the development of T regulatory cells, depends upon Kruppel-like factor 10 (KLF10) interacting with two antagonistic histone-modifying systems. We utilized chromatin immunoprecipitation, genome-integrated reporter assays, and functional domain KLF10 mutant proteins, to characterize reciprocal interactions between this transcription factor and either the Sin3-histone deacetylase complex or the histone acetyltransferase, p300/CBP-associated factor (PCAF). We characterize a Sin3-interacting repressor domain on the NH2 terminus of KLF10, which works to limit the activating function of this transcription factor. Indeed, inactivation of this Sin3-interacting domain renders KLF10 able to physically associate with PCAF as to induce FOXP3 gene transcription. We show that this biochemical data derived from studying our genome-integrated reporter cell system are recapitulated in primary murine lymphocytes. Collectively, these results advance our understanding of how a single transcription factor, namely KLF10, functions as a toggle to integrate antagonistic signals regulating FOXP3 and, thus, immune activation.
Keywords: FOXP3; KLF10; PCAF; Sin3; T regulatory cell.
Copyright © 2014 the American Physiological Society.
Figures











Similar articles
-
Polycomb antagonizes p300/CREB-binding protein-associated factor to silence FOXP3 in a Kruppel-like factor-dependent manner.J Biol Chem. 2012 Oct 5;287(41):34372-85. doi: 10.1074/jbc.M111.325332. Epub 2012 Aug 15. J Biol Chem. 2012. PMID: 22896699 Free PMC article.
-
Krüppel-like factor KLF10 deficiency predisposes to colitis through colonic macrophage dysregulation.Am J Physiol Gastrointest Liver Physiol. 2015 Dec 1;309(11):G900-9. doi: 10.1152/ajpgi.00309.2015. Epub 2015 Oct 15. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 26472224 Free PMC article.
-
Bone marrow-derived Kruppel-like factor 10 controls reendothelialization in response to arterial injury.Arterioscler Thromb Vasc Biol. 2013 Jul;33(7):1552-60. doi: 10.1161/ATVBAHA.112.300655. Epub 2013 May 16. Arterioscler Thromb Vasc Biol. 2013. PMID: 23685559 Free PMC article.
-
Co-repressor, co-activator and general transcription factor: the many faces of the Sin3 histone deacetylase (HDAC) complex.Biochem J. 2018 Dec 14;475(24):3921-3932. doi: 10.1042/BCJ20170314. Biochem J. 2018. PMID: 30552170 Free PMC article. Review.
-
Klf10 and Klf11 as mediators of TGF-beta superfamily signaling.Cell Tissue Res. 2012 Jan;347(1):65-72. doi: 10.1007/s00441-011-1186-6. Epub 2011 May 17. Cell Tissue Res. 2012. PMID: 21574058 Review.
Cited by
-
Forkhead transcription factor Fkh1: insights into functional regulatory domains crucial for recruitment of Sin3 histone deacetylase complex.Curr Genet. 2021 Jun;67(3):487-499. doi: 10.1007/s00294-021-01158-3. Epub 2021 Feb 26. Curr Genet. 2021. PMID: 33635403 Free PMC article.
-
Epigenetic Variability of CD4+CD25+ Tregs Contributes to the Pathogenesis of Autoimmune Diseases.Clin Rev Allergy Immunol. 2017 Apr;52(2):260-272. doi: 10.1007/s12016-016-8590-3. Clin Rev Allergy Immunol. 2017. PMID: 27687891 Review.
-
Kruppel-like Factors in Skeletal Physiology and Pathologies.Int J Mol Sci. 2022 Dec 2;23(23):15174. doi: 10.3390/ijms232315174. Int J Mol Sci. 2022. PMID: 36499521 Free PMC article. Review.
-
PCAF fine-tunes hepatic metabolic syndrome, inflammatory disease, and cancer.J Cell Mol Med. 2018 Dec;22(12):5787-5800. doi: 10.1111/jcmm.13877. Epub 2018 Sep 14. J Cell Mol Med. 2018. PMID: 30216660 Free PMC article. Review.
-
KLF10 Mediated Epigenetic Dysregulation of Epithelial CD40/CD154 Promotes Endometriosis.Biol Reprod. 2016 Sep;95(3):62. doi: 10.1095/biolreprod.116.140764. Epub 2016 Aug 3. Biol Reprod. 2016. PMID: 27488034 Free PMC article.
References
-
- Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441: 235–238, 2006 - PubMed
-
- Brubaker K, Cowley SM, Huang K, Loo L, Yochum GS, Ayer DE, Eisenman RN, Radhakrishnan I. Solution structure of the interacting domains of the Mad-Sin3 complex: implications for recruitment of a chromatin-modifying complex. Cell 103: 655–665, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous