

Regulation of Schistosoma mansoni development and reproduction by the mitogen-activated protein kinase signaling pathway
- PMID: 24945272
- PMCID: PMC4063740
- DOI: 10.1371/journal.pntd.0002949
Regulation of Schistosoma mansoni development and reproduction by the mitogen-activated protein kinase signaling pathway
Erratum in
- PLoS Negl Trop Dis. 2014 Jul;8(7):e3081
Abstract
Background: Protein kinases are proven targets for drug development with an increasing number of eukaryotic Protein Kinase (ePK) inhibitors now approved as drugs. Mitogen-activated protein kinase (MAPK) family members connect cell-surface receptors to regulatory targets within cells and influence a number of tissue-specific biological activities such as cell proliferation, differentiation and survival. However, the contributions of members of the MAPK pathway to schistosome development and survival are unclear.
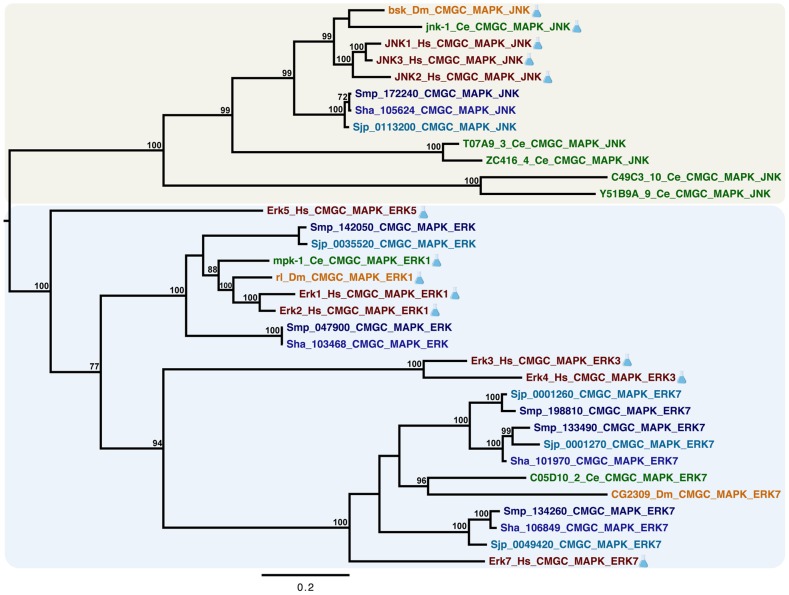
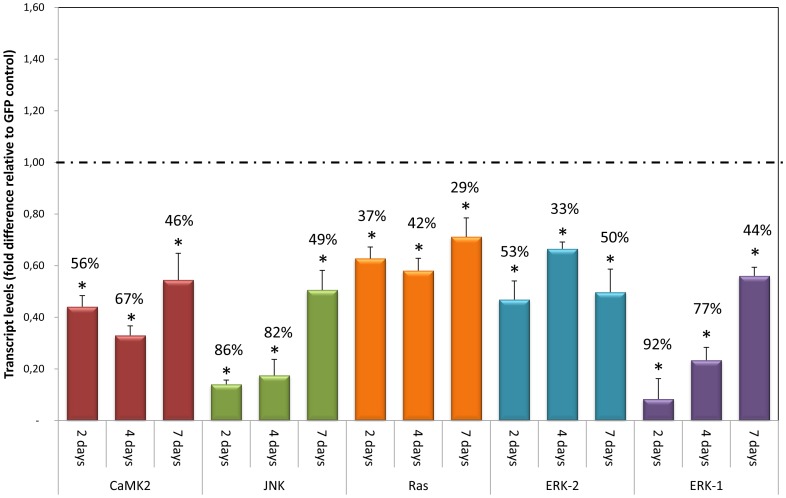
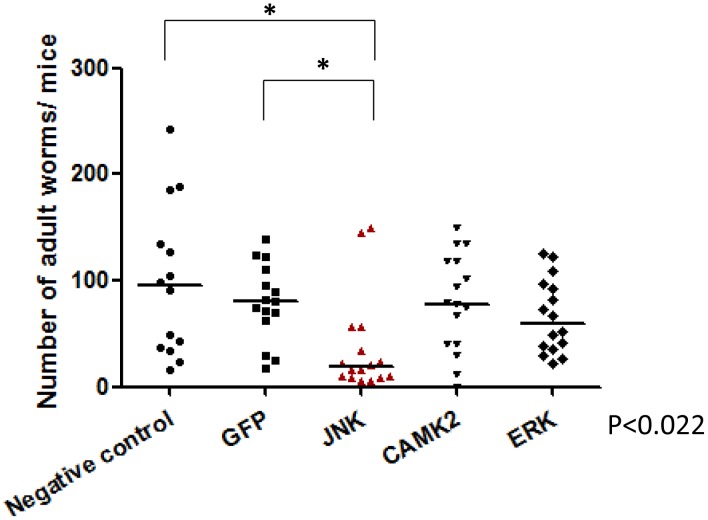
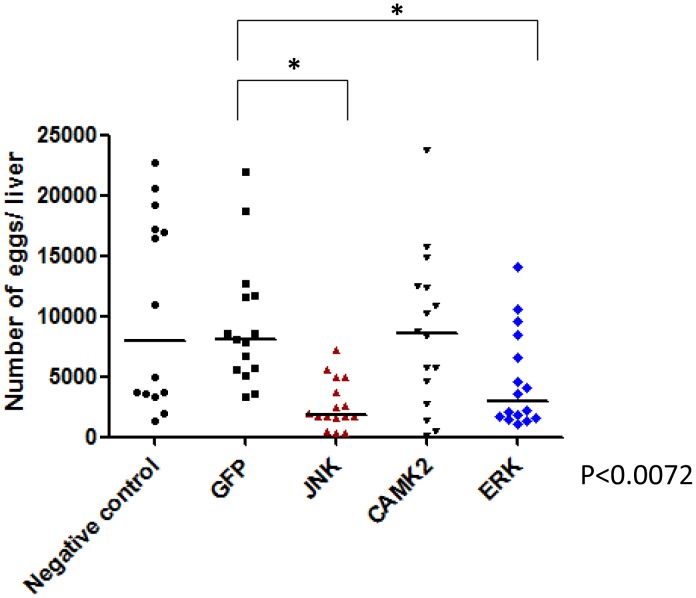
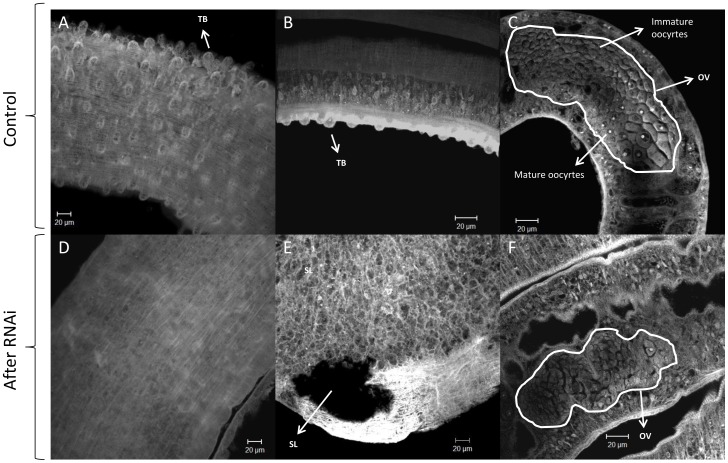
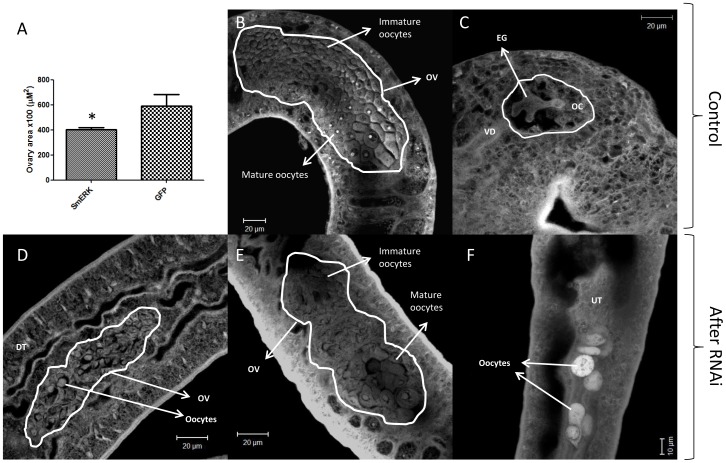
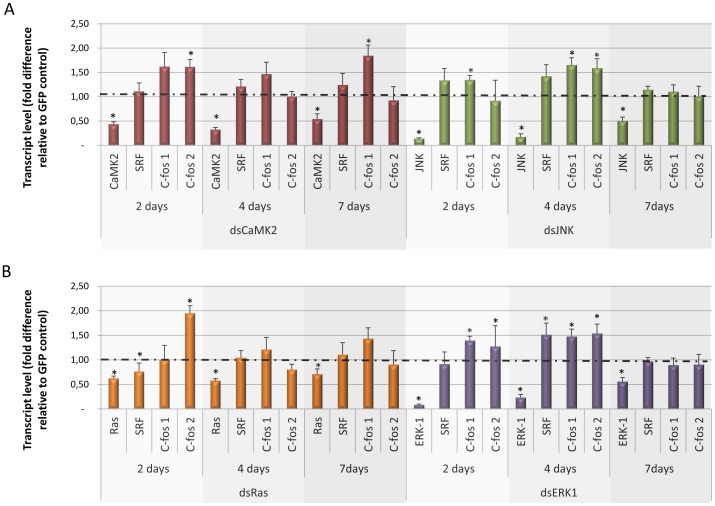
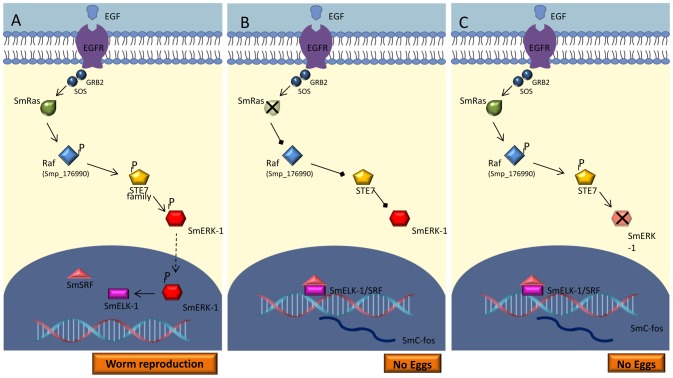
Methodology/principal findings: We employed RNA interference (RNAi) to elucidate the functional roles of five S. mansoni genes (SmCaMK2, SmJNK, SmERK1, SmERK2 and SmRas) involved in MAPK signaling pathway. Mice were injected with post-infective larvae (schistosomula) subsequent to RNAi and the development of adult worms observed. The data demonstrate that SmJNK participates in parasite maturation and survival of the parasites, whereas SmERK are involved in egg production as infected mice had significantly lower egg burdens with female worms presenting underdeveloped ovaries. Furthermore, it was shown that the c-fos transcription factor was overexpressed in parasites submitted to RNAi of SmERK1, SmJNK and SmCaMK2 indicating its putative involvement in gene regulation in this parasite's MAPK signaling cascade.
Conclusions: We conclude that MAPKs proteins play important roles in the parasite in vivo survival, being essential for normal development and successful survival and reproduction of the schistosome parasite. Moreover SmERK and SmJNK are potential targets for drug development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Allen JE, Maizels RM (2011) Diversity and dialogue in immunity to helminths. Nature Reviews Immunology 11: 375–388. - PubMed
-
- Pearce EJ, Sher A (1987) Mechanisms of immune evasion in schistosomiasis. Contrib Microbiol Immunol 8: 219–232. - PubMed
-
- Han ZG, Brindley PJ, Wang SY, Chen Z (2009) Schistosoma genomics: new perspectives on schistosome biology and host-parasite interaction. Annu.Rev.Genomics Hum.Genet 10: 211–240. - PubMed
-
- Wilkinson MG, Millar JB (2000) Control of the eukaryotic cell cycle by MAP kinase signaling pathways. FASEB J 14: 2147–2157. - PubMed
-
- Hazzalin CA, Mahadevan LC (2002) MAPK-regulated transcription: A continuously variable gene switch? Nature Reviews Molecular Cell Biology 3: 30–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
