

CPF-associated phosphatase activity opposes condensin-mediated chromosome condensation
- PMID: 24945319
- PMCID: PMC4063703
- DOI: 10.1371/journal.pgen.1004415
CPF-associated phosphatase activity opposes condensin-mediated chromosome condensation
Abstract
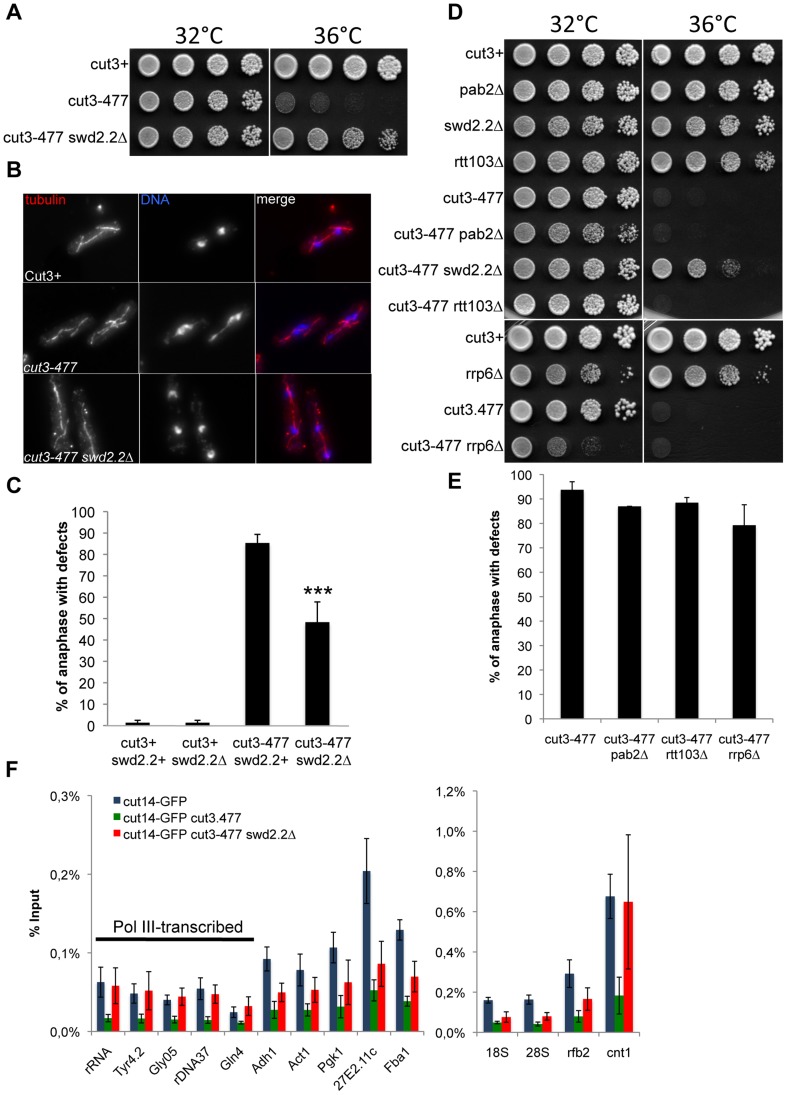
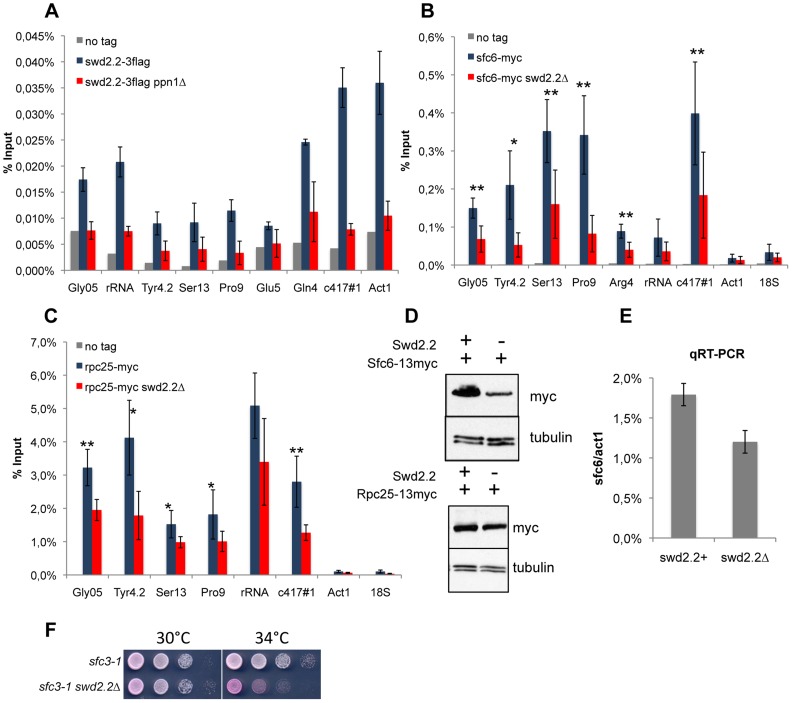
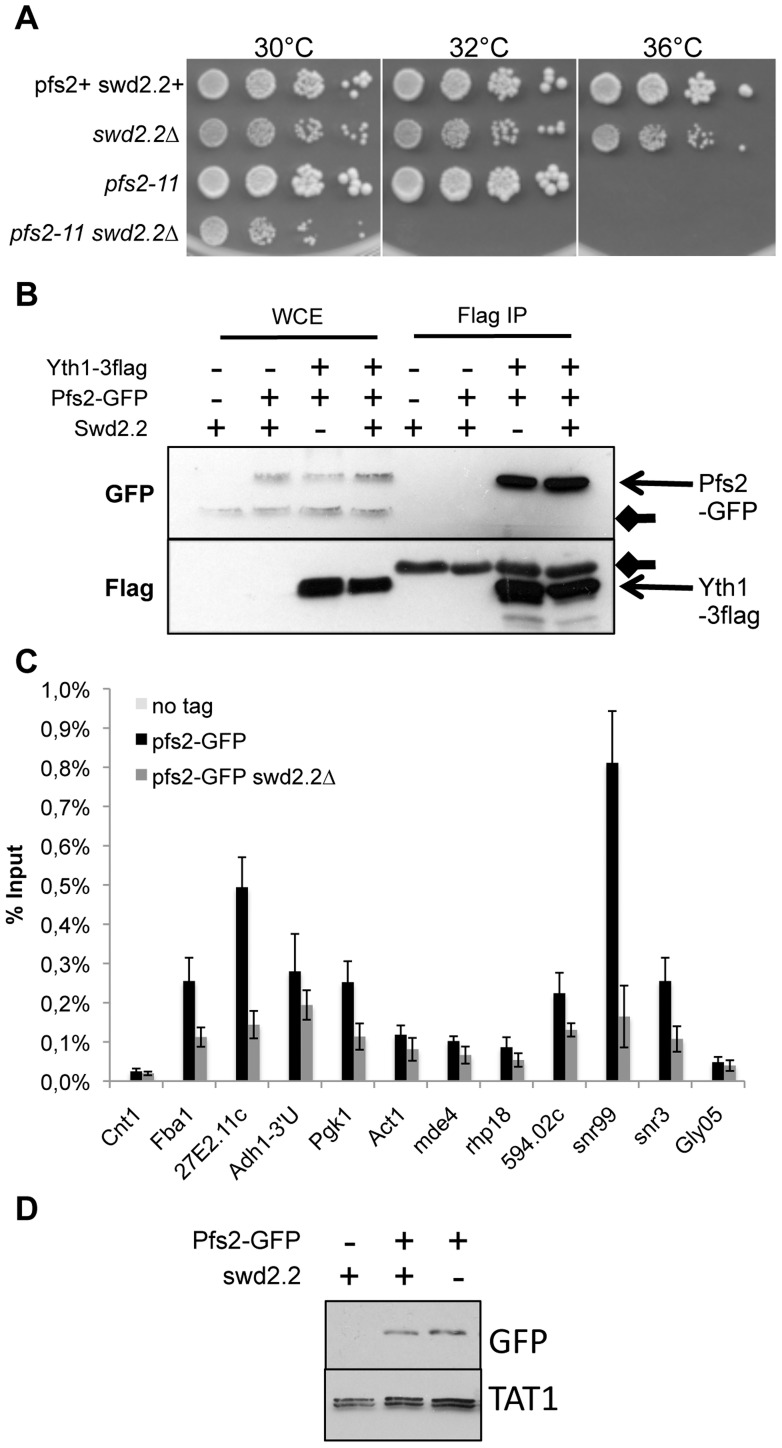
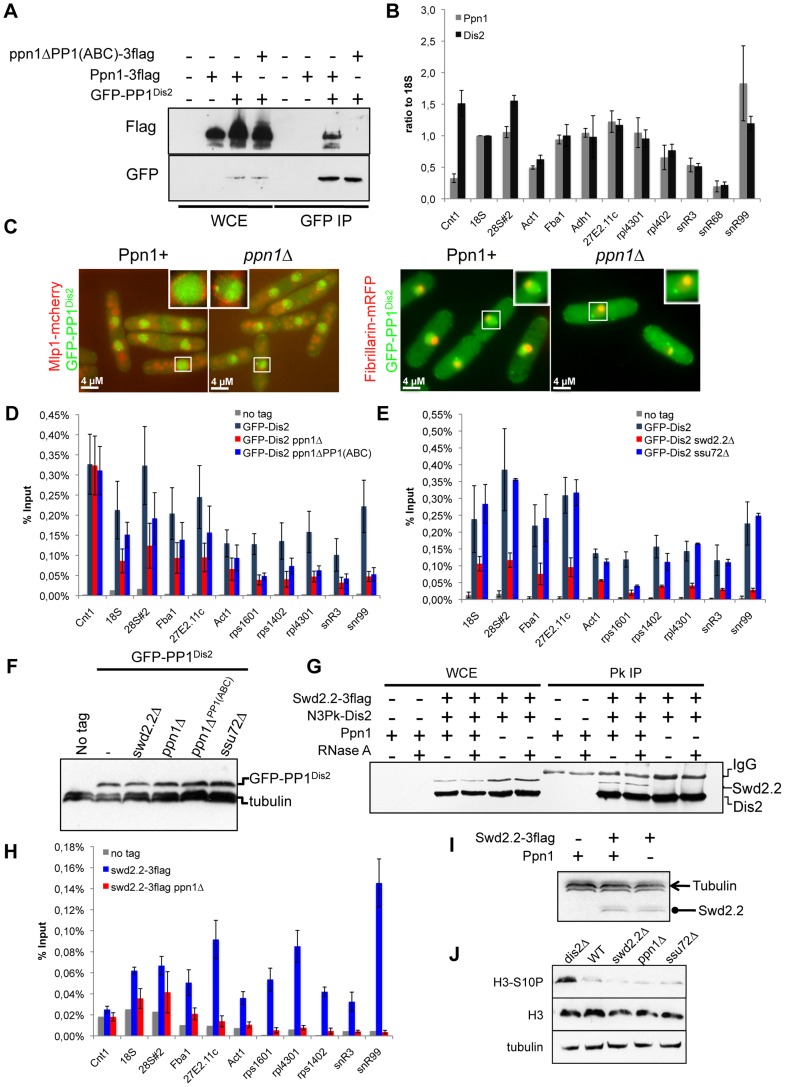
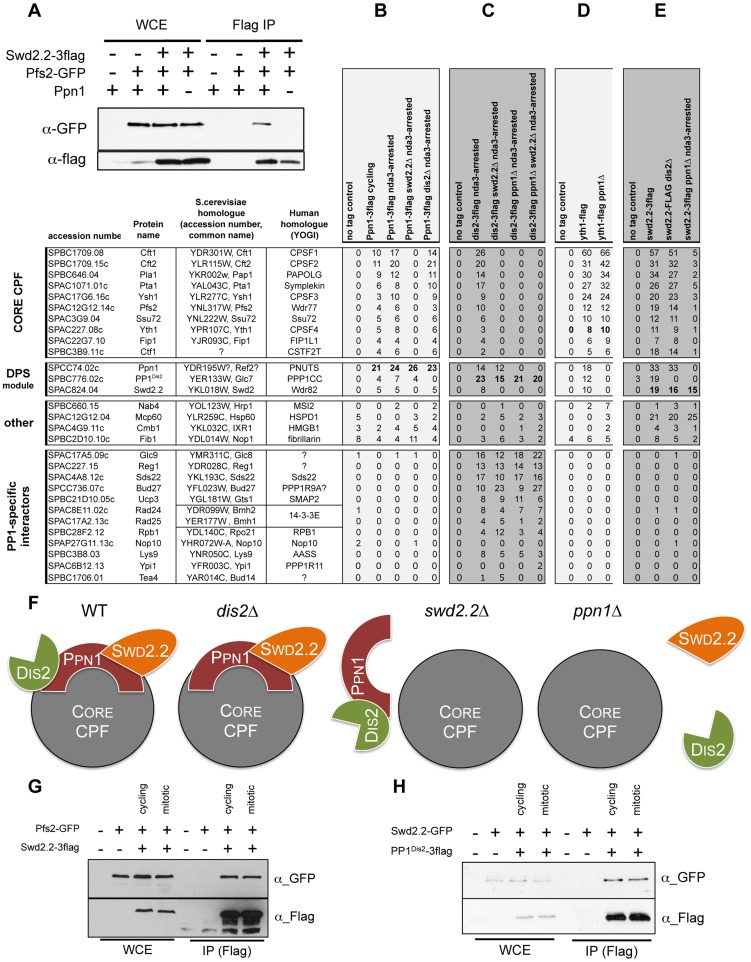
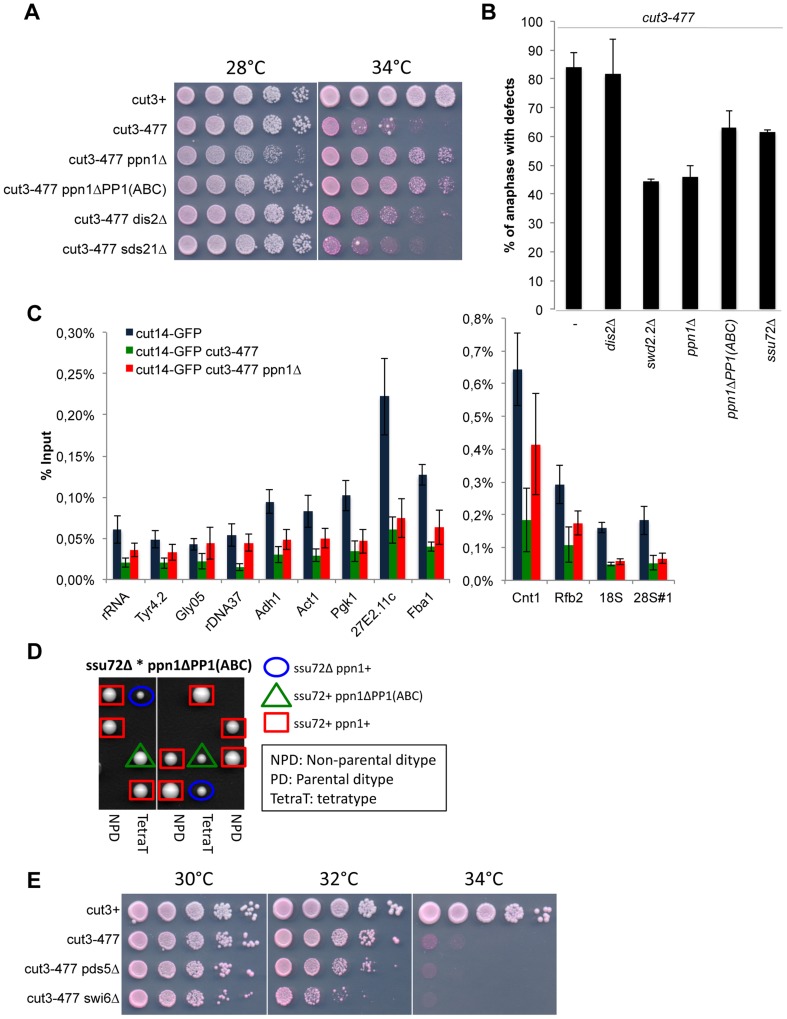
Functional links connecting gene transcription and condensin-mediated chromosome condensation have been established in species ranging from prokaryotes to vertebrates. However, the exact nature of these links remains misunderstood. Here we show in fission yeast that the 3' end RNA processing factor Swd2.2, a component of the Cleavage and Polyadenylation Factor (CPF), is a negative regulator of condensin-mediated chromosome condensation. Lack of Swd2.2 does not affect the assembly of the CPF but reduces its association with chromatin. This causes only limited, context-dependent effects on gene expression and transcription termination. However, CPF-associated Swd2.2 is required for the association of Protein Phosphatase 1 PP1(Dis2) with chromatin, through an interaction with Ppn1, a protein that we identify as the fission yeast homologue of vertebrate PNUTS. We demonstrate that Swd2.2, Ppn1 and PP1Dis2 form an independent module within the CPF, which provides an essential function in the absence of the CPF-associated Ssu72 phosphatase. We show that Ppn1 and Ssu72, like Swd2.2, are also negative regulators of condensin-mediated chromosome condensation. We conclude that Swd2.2 opposes condensin-mediated chromosome condensation by facilitating the function of the two CPF-associated phosphatases PP1 and Ssu72.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
