

The calcium-dependent protein kinase 3 of toxoplasma influences basal calcium levels and functions beyond egress as revealed by quantitative phosphoproteome analysis
- PMID: 24945436
- PMCID: PMC4063958
- DOI: 10.1371/journal.ppat.1004197
The calcium-dependent protein kinase 3 of toxoplasma influences basal calcium levels and functions beyond egress as revealed by quantitative phosphoproteome analysis
Abstract
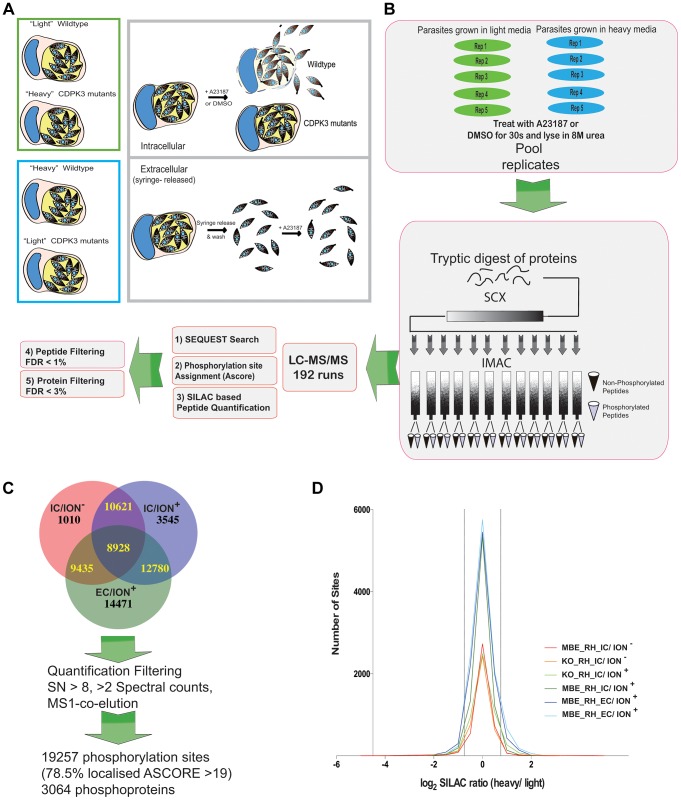
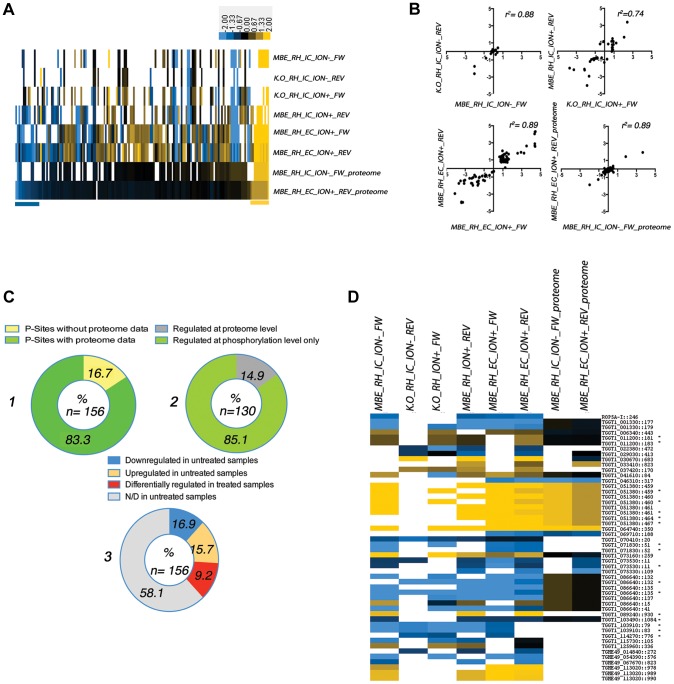
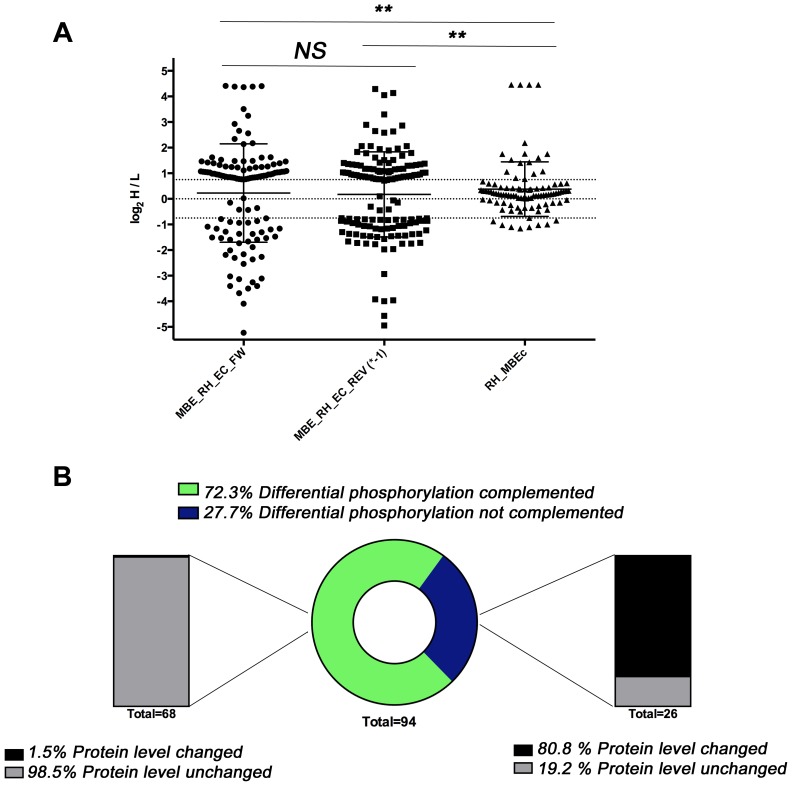
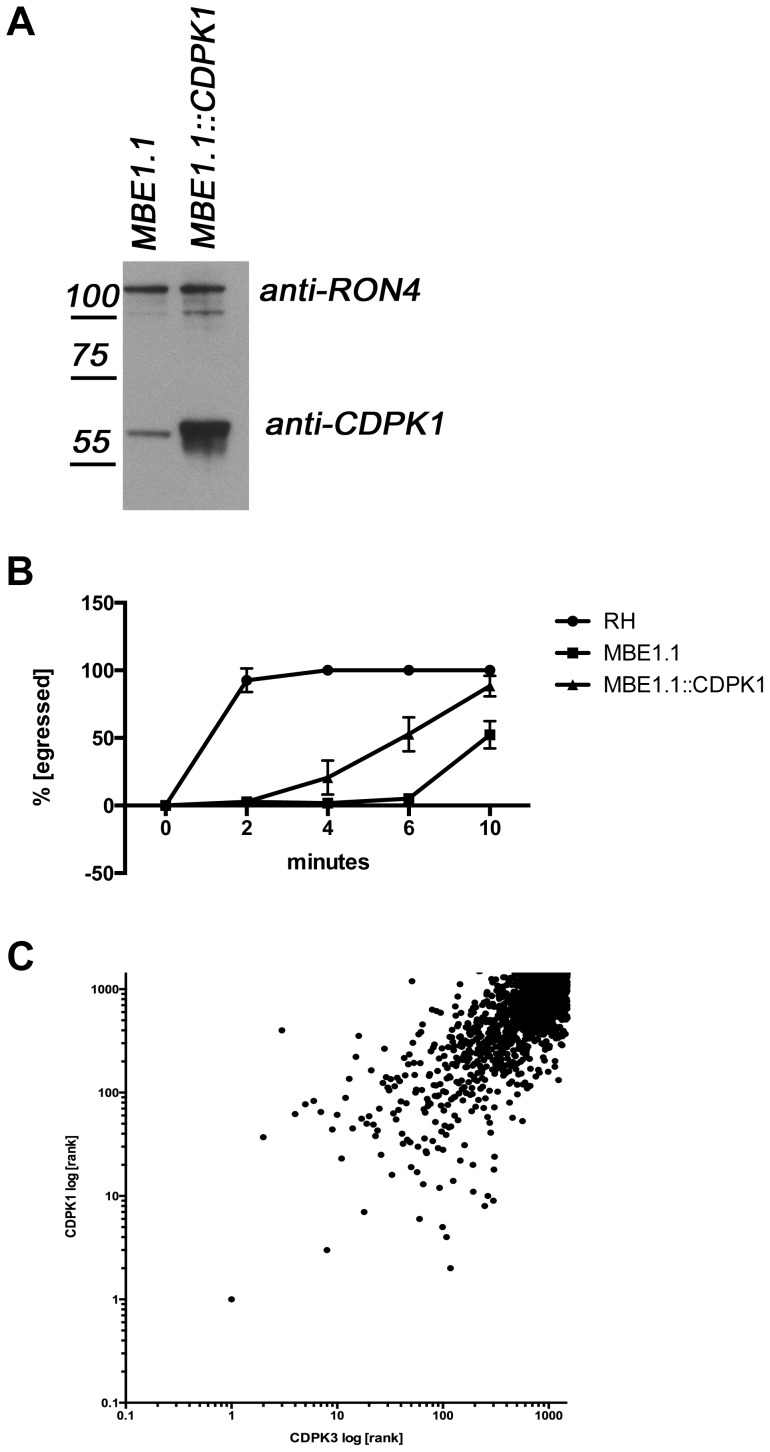
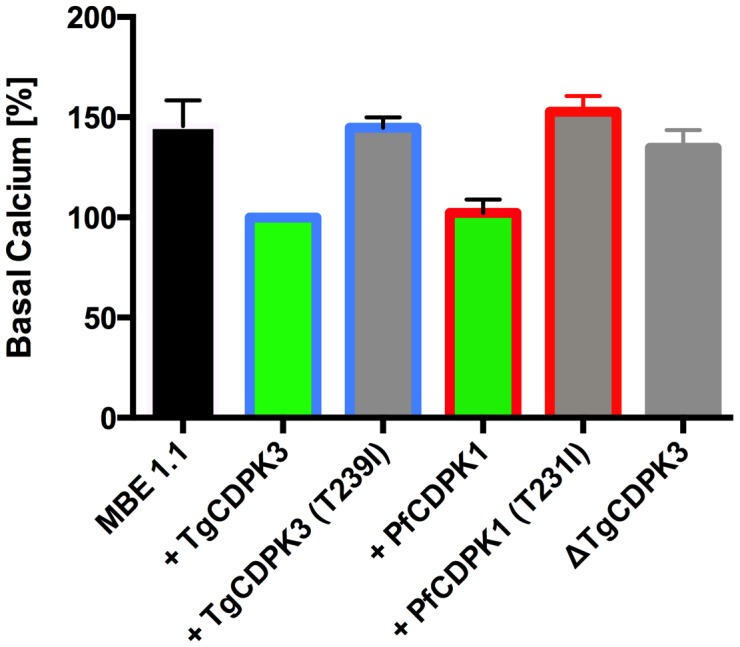
Calcium-dependent protein kinases (CDPKs) are conserved in plants and apicomplexan parasites. In Toxoplasma gondii, TgCDPK3 regulates parasite egress from the host cell in the presence of a calcium-ionophore. The targets and the pathways that the kinase controls, however, are not known. To identify pathways regulated by TgCDPK3, we measured relative phosphorylation site usage in wild type and TgCDPK3 mutant and knock-out parasites by quantitative mass-spectrometry using stable isotope-labeling with amino acids in cell culture (SILAC). This revealed known and novel phosphorylation events on proteins predicted to play a role in host-cell egress, but also a novel function of TgCDPK3 as an upstream regulator of other calcium-dependent signaling pathways, as we also identified proteins that are differentially phosphorylated prior to egress, including proteins important for ion-homeostasis and metabolism. This observation is supported by the observation that basal calcium levels are increased in parasites where TgCDPK3 has been inactivated. Most of the differential phosphorylation observed in CDPK3 mutants is rescued by complementation of the mutants with a wild type copy of TgCDPK3. Lastly, the TgCDPK3 mutants showed hyperphosphorylation of two targets of a related calcium-dependent kinase (TgCDPK1), as well as TgCDPK1 itself, indicating that this latter kinase appears to play a role downstream of TgCDPK3 function. Overexpression of TgCDPK1 partially rescues the egress phenotype of the TgCDPK3 mutants, reinforcing this conclusion. These results show that TgCDPK3 plays a pivotal role in regulating tachyzoite functions including, but not limited to, egress.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Billker O, Dechamps S, Tewari R, Wenig G, Franke-Fayard B, et al. (2004) Calcium and a calcium-dependent protein kinase regulate gamete formation and mosquito transmission in a malaria parasite. Cell 117: 503–514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
