

Co-ordinated regulation of the extracytoplasmic stress factor, sigmaE, with other Escherichia coli sigma factors by (p)ppGpp and DksA may be achieved by specific regulation of individual holoenzymes
- PMID: 24946009
- PMCID: PMC4497380
- DOI: 10.1111/mmi.12674
Co-ordinated regulation of the extracytoplasmic stress factor, sigmaE, with other Escherichia coli sigma factors by (p)ppGpp and DksA may be achieved by specific regulation of individual holoenzymes
Abstract
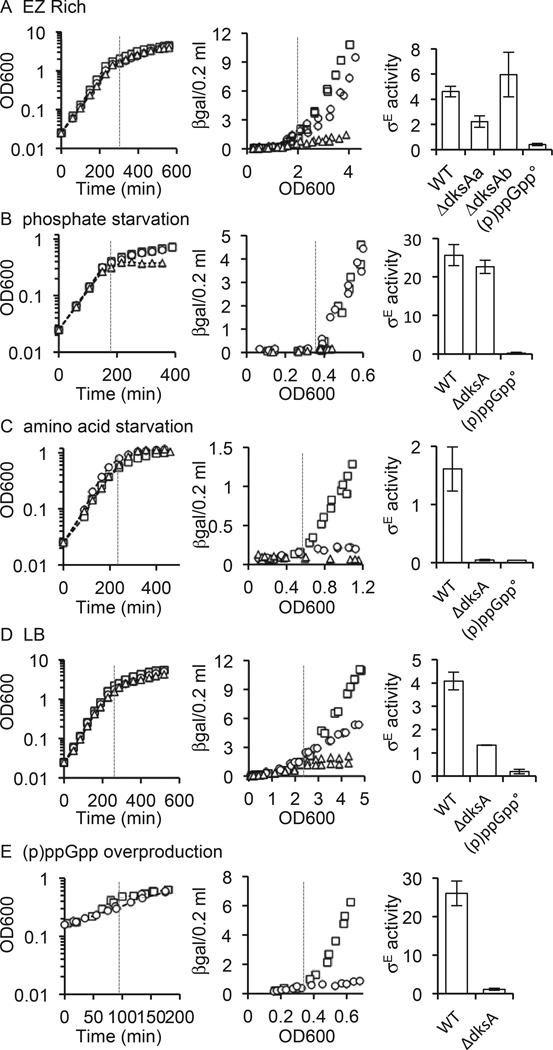
The E. coli alternative sigma factor, σ(E) , transcribes genes required to maintain the cell envelope and is activated by conditions that destabilize the envelope. σ(E) is also activated during entry into stationary phase in the absence of envelope stress by the alarmone (p)ppGpp. (p)ppGpp controls a large regulatory network, reducing expression of σ(70) -dependent genes required for rapid growth and activating σ(70) -dependent and alternative sigma factor-dependent genes required for stress survival. The DksA protein often potentiates the effects of (p)ppGpp. Here we examine regulation of σ(E) by (p)ppGpp and DksA following starvation for nutrients. We find that (p)ppGpp is required for increased σ(E) activity under all conditions tested, but the requirement for DksA varies. DksA is required during amino acid starvation, but is dispensable during phosphate starvation. In contrast, regulation of σ(S) is (p)ppGpp- and DksA-dependent under all conditions tested, while negative regulation of σ(70) is DksA- but not (p)ppGpp-dependent during phosphate starvation, yet requires both factors during amino acid starvation. These findings suggest that the mechanism of transcriptional regulation by (p)ppGpp and/or DksA cannot yet be explained by a unifying model and is specific to individual promoters, individual holoenzymes, and specific starvation conditions.
© 2014 John Wiley & Sons Ltd.
Figures







References
-
- Barchinger SE, Ades SE. Regulated proteolysis: control of the Escherichia coli σ(E)-dependent cell envelope stress response. Subcell Biochem. 2013;66:129–160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
