

Genome-wide identification and functional analysis of Apobec-1-mediated C-to-U RNA editing in mouse small intestine and liver
- PMID: 24946870
- PMCID: PMC4197816
- DOI: 10.1186/gb-2014-15-6-r79
Genome-wide identification and functional analysis of Apobec-1-mediated C-to-U RNA editing in mouse small intestine and liver
Abstract
Background: RNA editing encompasses a post-transcriptional process in which the genomically templated sequence is enzymatically altered and introduces a modified base into the edited transcript. Mammalian C-to-U RNA editing represents a distinct subtype of base modification, whose prototype is intestinal apolipoprotein B mRNA, mediated by the catalytic deaminase Apobec-1. However, the genome-wide identification, tissue-specificity and functional implications of Apobec-1-mediated C-to-U RNA editing remain incompletely explored.
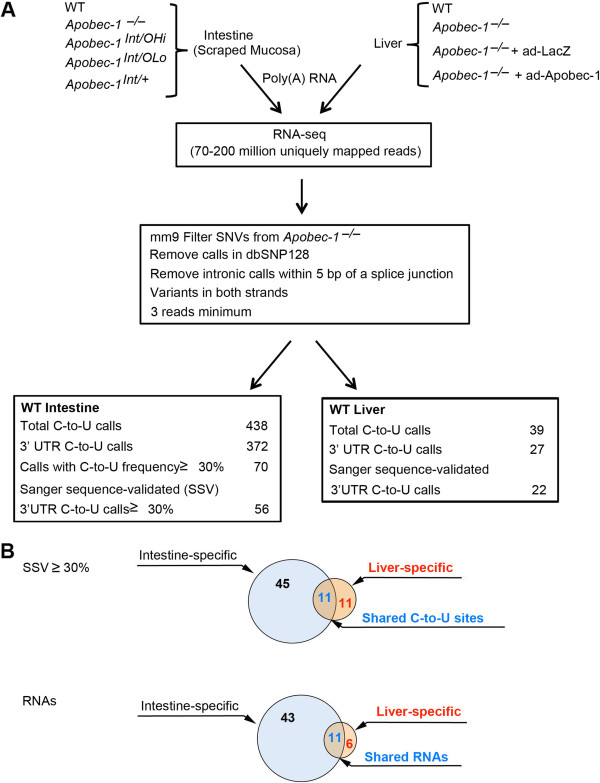
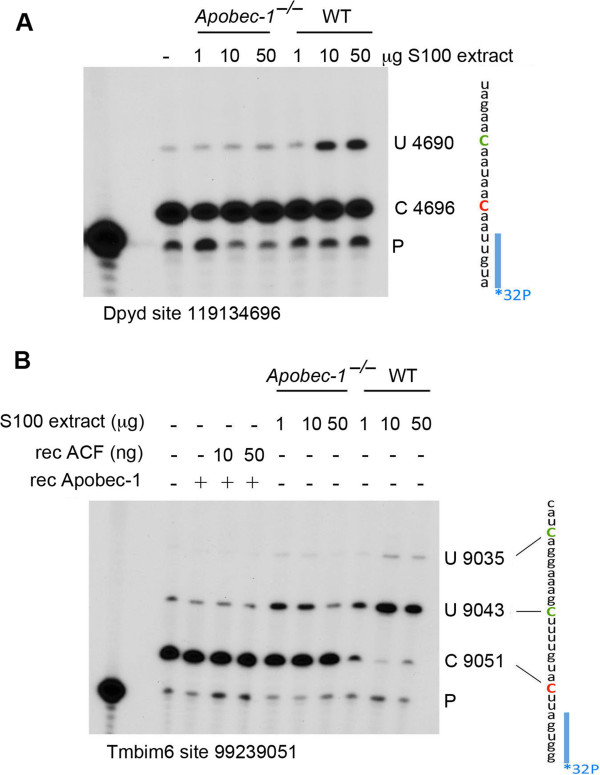
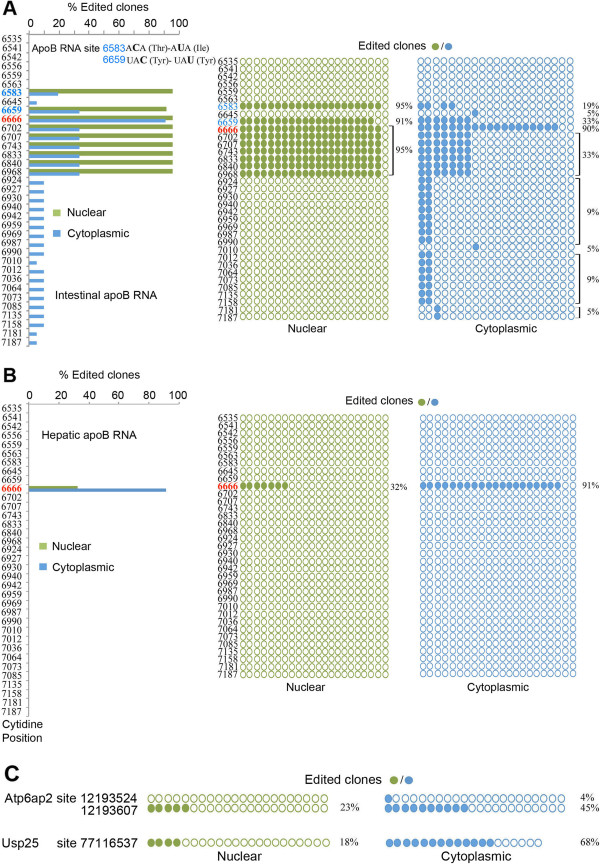
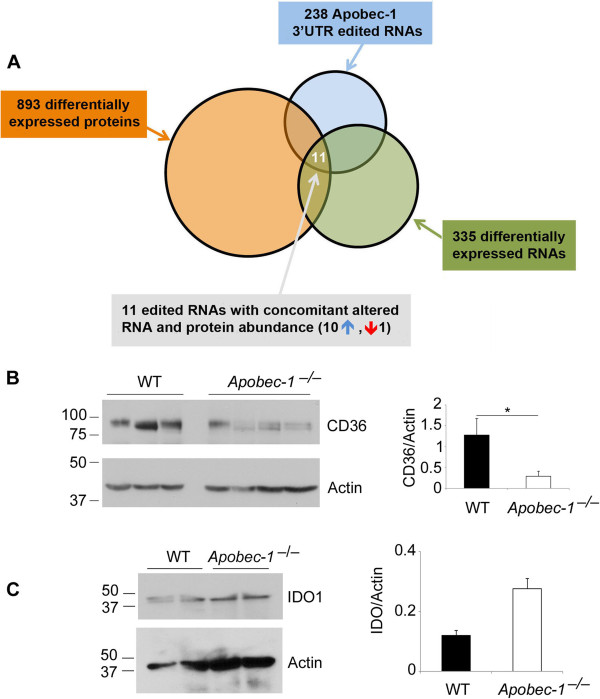
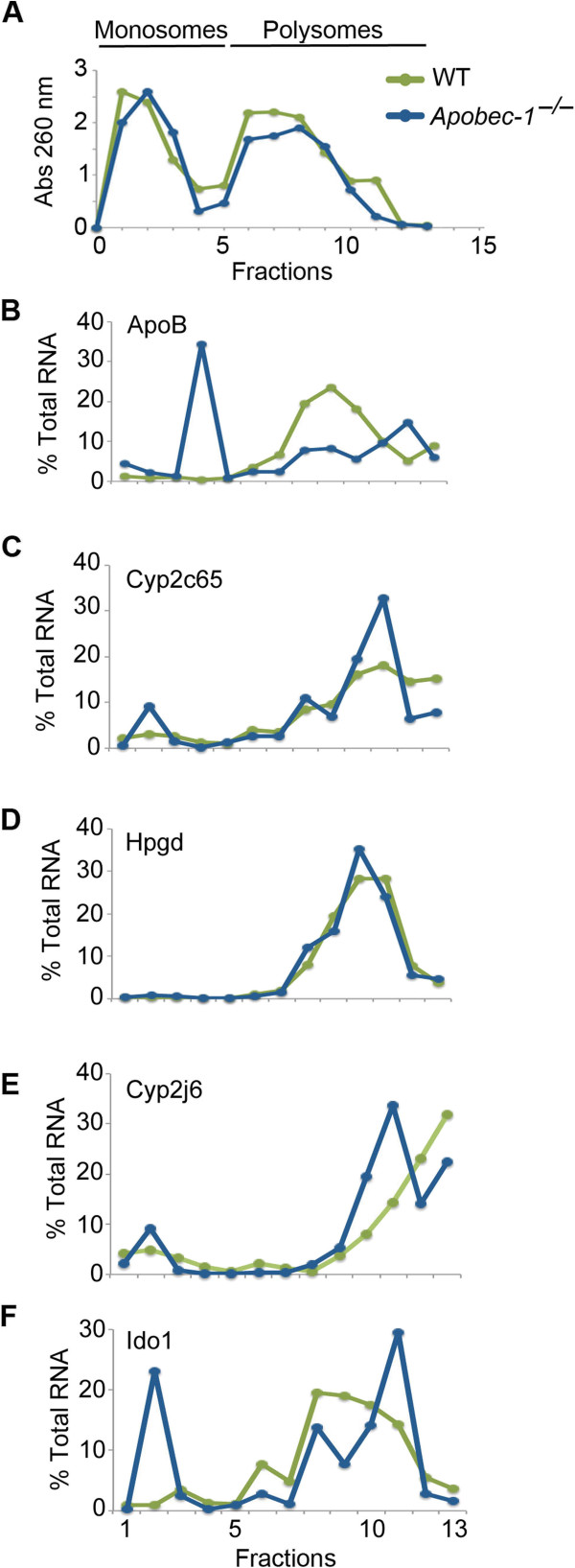
Results: Deep sequencing, data filtering and Sanger-sequence validation of intestinal and hepatic RNA from wild-type and Apobec-1-deficient mice revealed 56 novel editing sites in 54 intestinal mRNAs and 22 novel sites in 17 liver mRNAs, all within 3' untranslated regions. Eleven of 17 liver RNAs shared editing sites with intestinal RNAs, while 6 sites are unique to liver. Changes in RNA editing lead to corresponding changes in intestinal mRNA and protein levels for 11 genes. Analysis of RNA editing in vivo following tissue-specific Apobec-1 adenoviral or transgenic Apobec-1 overexpression reveals that a subset of targets identified in wild-type mice are restored in Apobec-1-deficient mouse intestine and liver following Apobec-1 rescue. We find distinctive polysome profiles for several RNA editing targets and demonstrate novel exonic editing sites in nuclear preparations from intestine but not hepatic apolipoprotein B RNA. RNA editing is validated using cell-free extracts from wild-type but not Apobec-1-deficient mice, demonstrating that Apobec-1 is required.
Conclusions: These studies define selective, tissue-specific targets of Apobec-1-dependent RNA editing and show the functional consequences of editing are both transcript- and tissue-specific.
Figures
References
-
- Peng Z, Cheng Y, Tan BC, Kang L, Tian Z, Zhu Y, Zhang W, Liang Y, Hu X, Tan X, Guo J, Dong Z, Liang Y, Bao L, Wang J. Comprehensive analysis of RNA-Seq data reveals extensive RNA editing in a human transcriptome. Nat Biotechnol. 2012;30:253–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA075056/CA/NCI NIH HHS/United States
- P30 DK-52574/DK/NIDDK NIH HHS/United States
- DP1 HD075624/HD/NICHD NIH HHS/United States
- CA75056/CA/NCI NIH HHS/United States
- P30 CA91842/CA/NCI NIH HHS/United States
- DK-56260/DK/NIDDK NIH HHS/United States
- U54HG006998/HG/NHGRI NIH HHS/United States
- DP1 OD006911/OD/NIH HHS/United States
- R01 HL038180/HL/NHLBI NIH HHS/United States
- UL1TR000448/TR/NCATS NIH HHS/United States
- HL-38180/HL/NHLBI NIH HHS/United States
- R01 DK056260/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
