

Evidence supporting the 19 β-strand model for Tom40 from cysteine scanning and protease site accessibility studies
- PMID: 24947507
- PMCID: PMC4118123
- DOI: 10.1074/jbc.M114.578765
Evidence supporting the 19 β-strand model for Tom40 from cysteine scanning and protease site accessibility studies
Abstract
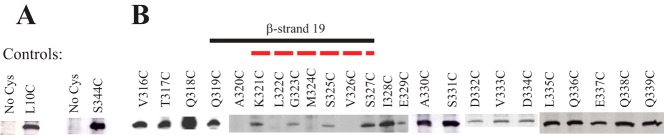
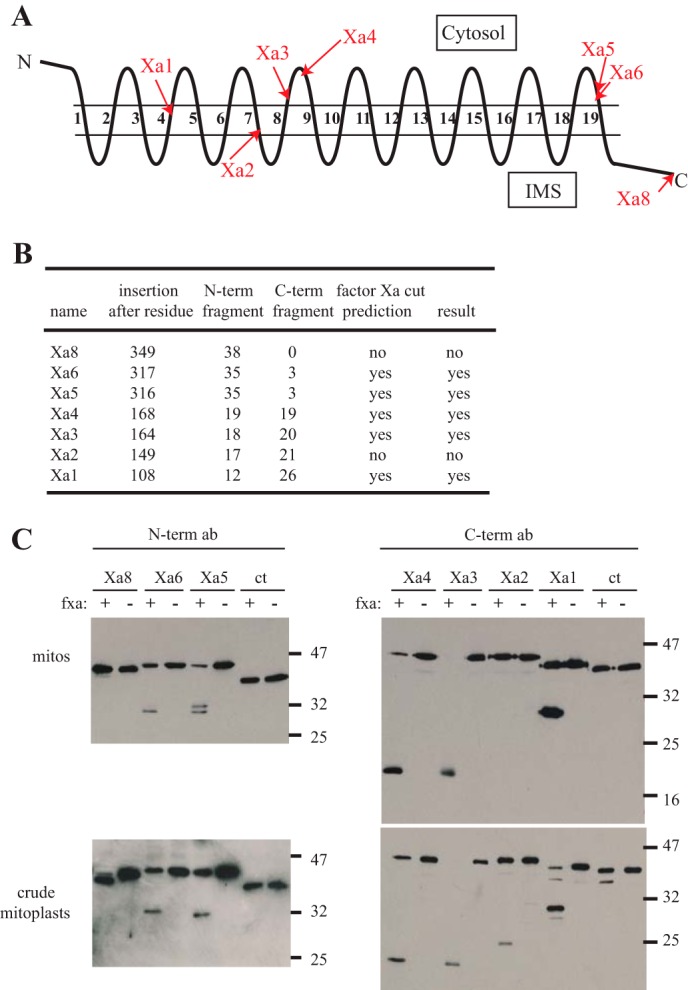
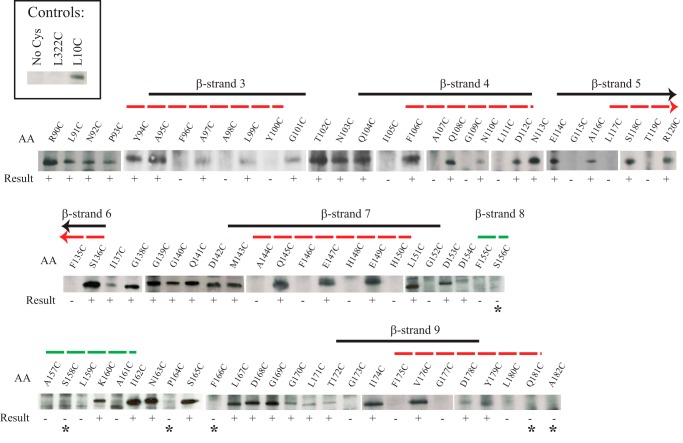
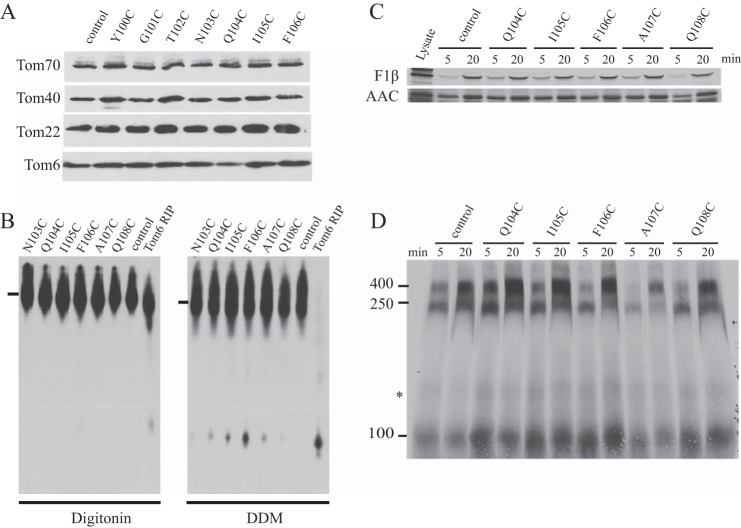
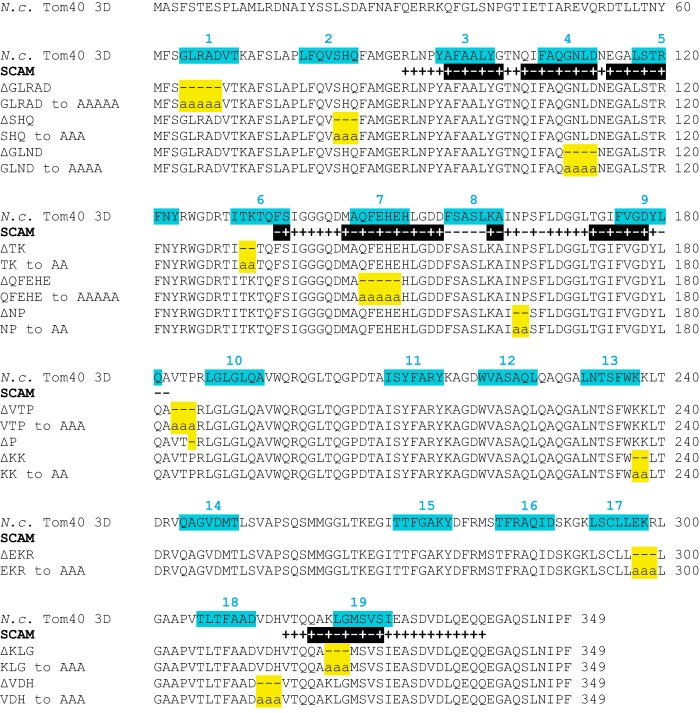
Most proteins found in mitochondria are translated in the cytosol and enter the organelle via the TOM complex (translocase of the outer mitochondrial membrane). Tom40 is the pore forming component of the complex. Although the three-dimensional structure of Tom40 has not been determined, the structure of porin, a related protein, has been shown to be a β-barrel containing 19 membrane spanning β-strands and an N-terminal α-helical region. The evolutionary relationship between the two proteins has allowed modeling of Tom40 into a similar structure by several laboratories. However, it has been suggested that the 19-strand porin structure does not represent the native form of the protein. If true, modeling of Tom40 based on the porin structure would also be invalid. We have used substituted cysteine accessibility mapping to identify several potential β-strands in the Tom40 protein in isolated mitochondria. These data, together with protease accessibility studies, support the 19 β-strand model for Tom40 with the C-terminal end of the protein localized to the intermembrane space.
Keywords: Membrane Protein; Mitochondria; Neurospora; Protein Structure; SCAM; Tom40; Transmembrane Domain; β-Barrel Protein.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

Similar articles
-
Structural elements of the mitochondrial preprotein-conducting channel Tom40 dissolved by bioinformatics and mass spectrometry.Biochim Biophys Acta. 2011 Dec;1807(12):1647-57. doi: 10.1016/j.bbabio.2011.08.006. Epub 2011 Aug 23. Biochim Biophys Acta. 2011. PMID: 21888892
-
Characterization of Neurospora crassa Tom40-deficient mutants and effect of specific mutations on Tom40 assembly.J Biol Chem. 2003 Jan 10;278(2):765-75. doi: 10.1074/jbc.M208083200. Epub 2002 Oct 23. J Biol Chem. 2003. PMID: 12399467
-
The Tim8-Tim13 complex of Neurospora crassa functions in the assembly of proteins into both mitochondrial membranes.J Biol Chem. 2004 Mar 26;279(13):12396-405. doi: 10.1074/jbc.M313037200. Epub 2004 Jan 13. J Biol Chem. 2004. PMID: 14722057
-
Protein import into mitochondria of Neurospora crassa.Fungal Genet Biol. 2002 Jul;36(2):85-90. doi: 10.1016/S1087-1845(02)00018-X. Fungal Genet Biol. 2002. PMID: 12081461 Review.
-
How does the TOM complex mediate insertion of precursor proteins into the mitochondrial outer membrane?J Cell Biol. 2005 Nov 7;171(3):419-23. doi: 10.1083/jcb.200507147. Epub 2005 Oct 31. J Cell Biol. 2005. PMID: 16260501 Free PMC article. Review.
Cited by
-
Role of the Mitochondrial Protein Import Machinery and Protein Processing in Heart Disease.Front Cardiovasc Med. 2021 Sep 28;8:749756. doi: 10.3389/fcvm.2021.749756. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34651031 Free PMC article. Review.
-
Cell biology: Architecture of a protein entry gate.Nature. 2015 Dec 10;528(7581):201-2. doi: 10.1038/nature16318. Epub 2015 Nov 25. Nature. 2015. PMID: 26605527 No abstract available.
-
Cryo-EM structure of the mitochondrial protein-import channel TOM complex at near-atomic resolution.Nat Struct Mol Biol. 2019 Dec;26(12):1158-1166. doi: 10.1038/s41594-019-0339-2. Epub 2019 Nov 18. Nat Struct Mol Biol. 2019. PMID: 31740857 Free PMC article.
-
Translocase of the Outer Mitochondrial Membrane 40 Is Required for Mitochondrial Biogenesis and Embryo Development in Arabidopsis.Front Plant Sci. 2019 Apr 2;10:389. doi: 10.3389/fpls.2019.00389. eCollection 2019. Front Plant Sci. 2019. PMID: 31001303 Free PMC article.
-
Ten Years of High Resolution Structural Research on the Voltage Dependent Anion Channel (VDAC)-Recent Developments and Future Directions.Front Physiol. 2018 Mar 7;9:108. doi: 10.3389/fphys.2018.00108. eCollection 2018. Front Physiol. 2018. PMID: 29563878 Free PMC article. Review.
References
-
- Neupert W., Herrmann J. (2007) Translocation of proteins into mitochondria. Annu. Rev. Biochem. 76, 723–749 - PubMed
-
- Endo T., Yamano K. (2009) Multiple pathways for mitochondrial protein traffic. Biol. Chem. 390, 723–730 - PubMed
-
- Schmidt O., Pfanner N., Meisinger C. (2010) Mitochondrial protein import: from proteomics to functional mechanisms. Nat. Rev. Mol. Cell Biol. 11, 655–667 - PubMed
-
- Becker T., Böttinger L., Pfanner N. (2012) Mitochondrial protein import: from transport pathways to an integrated network. Trends Biochem. Sci. 37, 85–91 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
