

Determination of the Structure and Catalytic Mechanism of Sorghum bicolor Caffeic Acid O-Methyltransferase and the Structural Impact of Three brown midrib12 Mutations
- PMID: 24948836
- PMCID: PMC4119030
- DOI: 10.1104/pp.114.241729
Determination of the Structure and Catalytic Mechanism of Sorghum bicolor Caffeic Acid O-Methyltransferase and the Structural Impact of Three brown midrib12 Mutations
Abstract
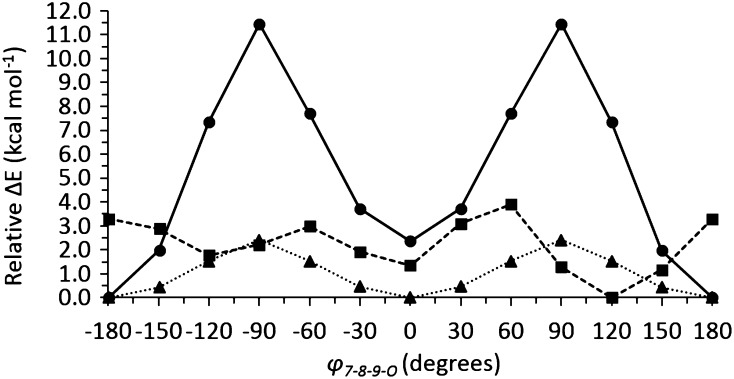
Using S-adenosyl-methionine as the methyl donor, caffeic acid O-methyltransferase from sorghum (Sorghum bicolor; SbCOMT) methylates the 5-hydroxyl group of its preferred substrate, 5-hydroxyconiferaldehyde. In order to determine the mechanism of SbCOMT and understand the observed reduction in the lignin syringyl-to-guaiacyl ratio of three brown midrib12 mutants that carry COMT gene missense mutations, we determined the apo-form and S-adenosyl-methionine binary complex SbCOMT crystal structures and established the ternary complex structure with 5-hydroxyconiferaldehyde by molecular modeling. These structures revealed many features shared with monocot ryegrass (Lolium perenne) and dicot alfalfa (Medicago sativa) COMTs. SbCOMT steady-state kinetic and calorimetric data suggest a random bi-bi mechanism. Based on our structural, kinetic, and thermodynamic results, we propose that the observed reactivity hierarchy among 4,5-dihydroxy-3-methoxycinnamyl (and 3,4-dihydroxycinnamyl) aldehyde, alcohol, and acid substrates arises from the ability of the aldehyde to stabilize the anionic intermediate that results from deprotonation of the 5-hydroxyl group by histidine-267. Additionally, despite the presence of other phenylpropanoid substrates in vivo, sinapaldehyde is the preferential product, as demonstrated by its low Km for 5-hydroxyconiferaldehyde. Unlike its acid and alcohol substrates, the aldehydes exhibit product inhibition, and we propose that this is due to nonproductive binding of the S-cis-form of the aldehydes inhibiting productive binding of the S-trans-form. The S-cis-aldehydes most likely act only as inhibitors, because the high rotational energy barrier around the 2-propenyl bond prevents S-trans-conversion, unlike alcohol substrates, whose low 2-propenyl bond rotational energy barrier enables rapid S-cis/S-trans-interconversion.
© 2014 American Society of Plant Biologists. All Rights Reserved.
Figures










References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC. (2002) PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr 58: 1948–1954 - PubMed
-
- Atanassova R, Favet N, Martz F, Chabbert B, Tollier M, Monties B, Fritig B, Legrand M. (1995) Altered lignin composition in transgenic tobacco expressing O-methyltransferase sequences in sense and antisense orientation. Plant J 8: 465–477
-
- Berlin A, Balakshin M, Gilkes N, Kadla J, Maximenko V, Kubo S, Saddler J. (2006) Inhibition of cellulase, xylanase and β-glucosidase activities by softwood lignin preparations. J Biotechnol 125: 198–209 - PubMed
-
- Bout S, Vermerris W. (2003) A candidate-gene approach to clone the sorghum Brown midrib gene encoding caffeic acid O-methyltransferase. Mol Genet Genomics 269: 205–214 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
