

Programmed translational readthrough generates antiangiogenic VEGF-Ax
- PMID: 24949972
- PMCID: PMC4113015
- DOI: 10.1016/j.cell.2014.04.033
Programmed translational readthrough generates antiangiogenic VEGF-Ax
Abstract
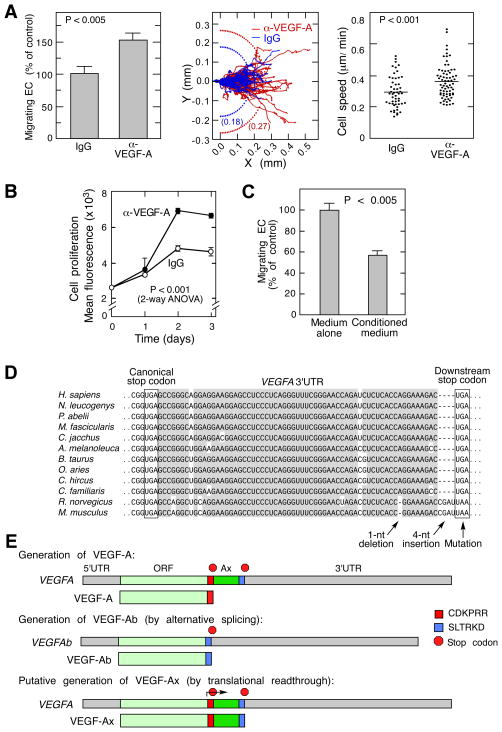
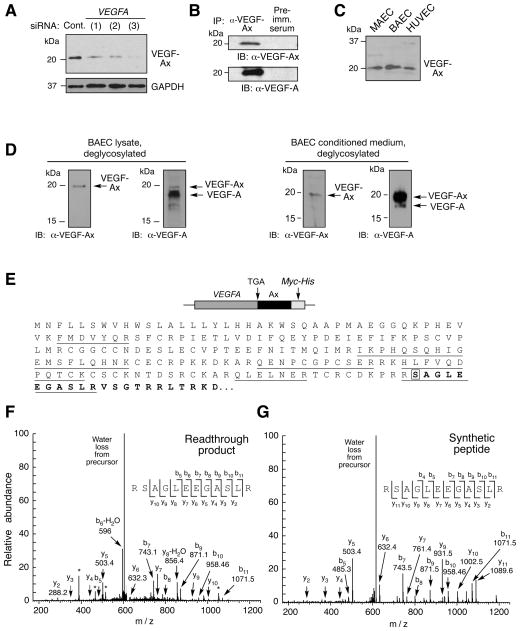
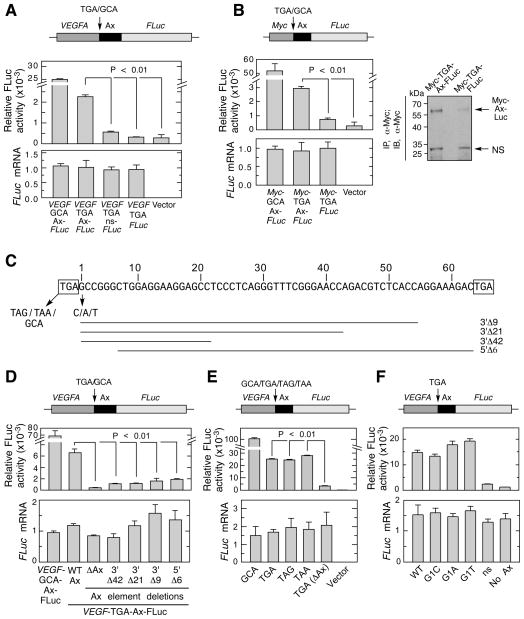
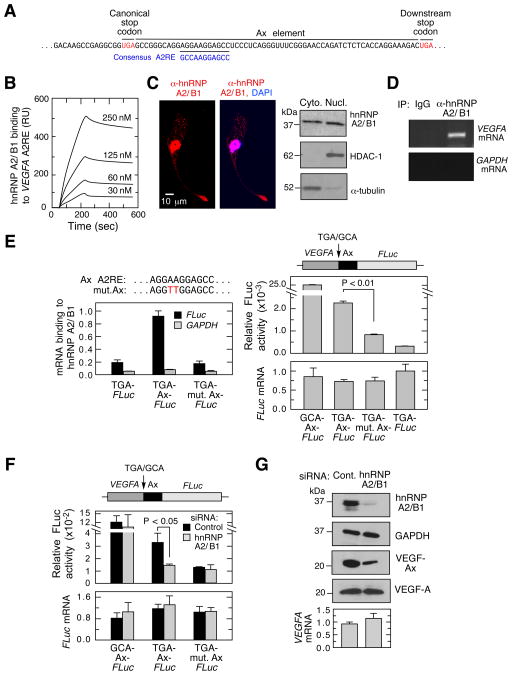
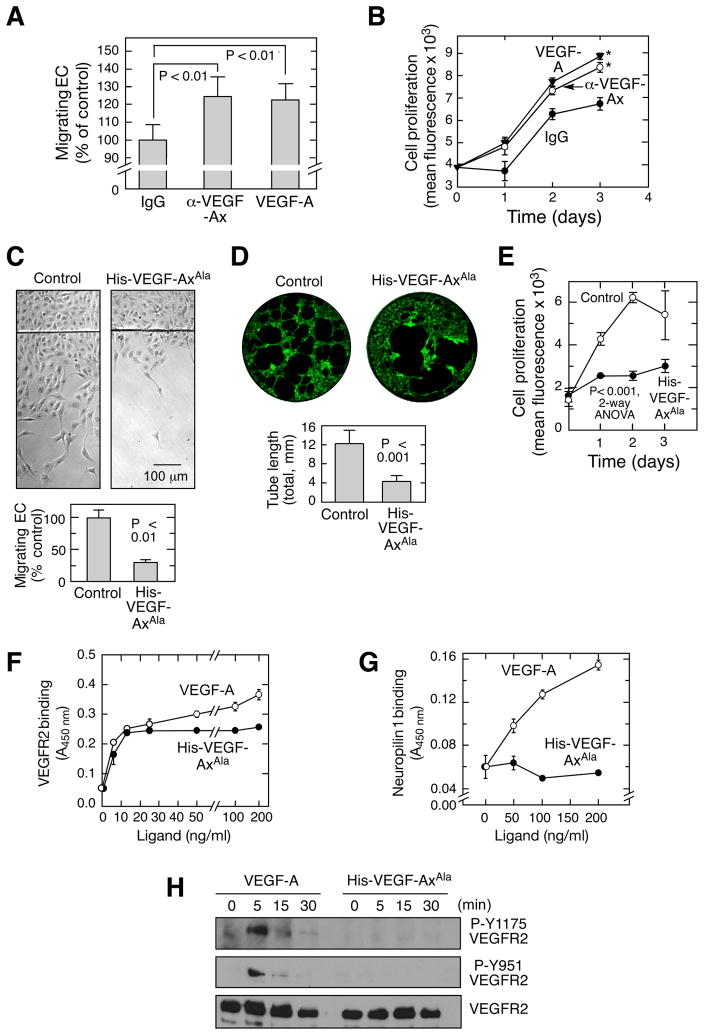
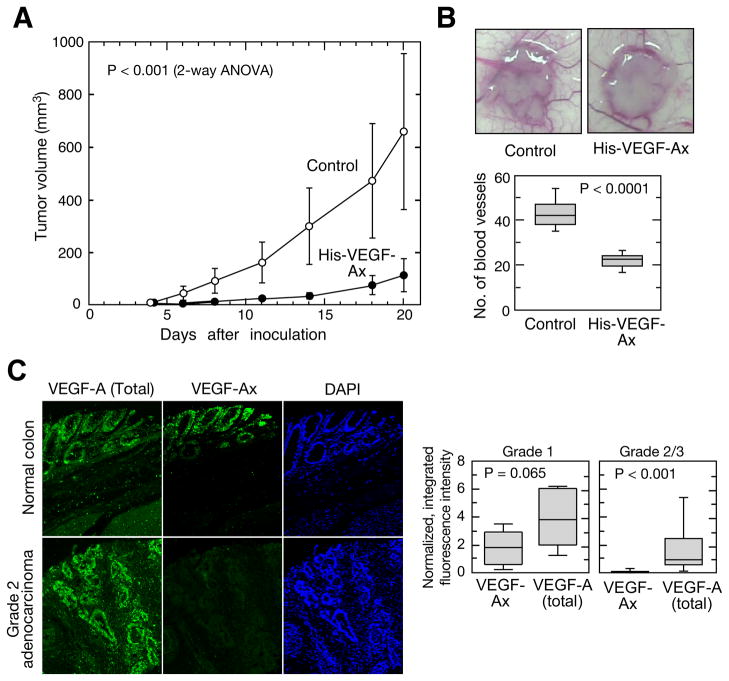
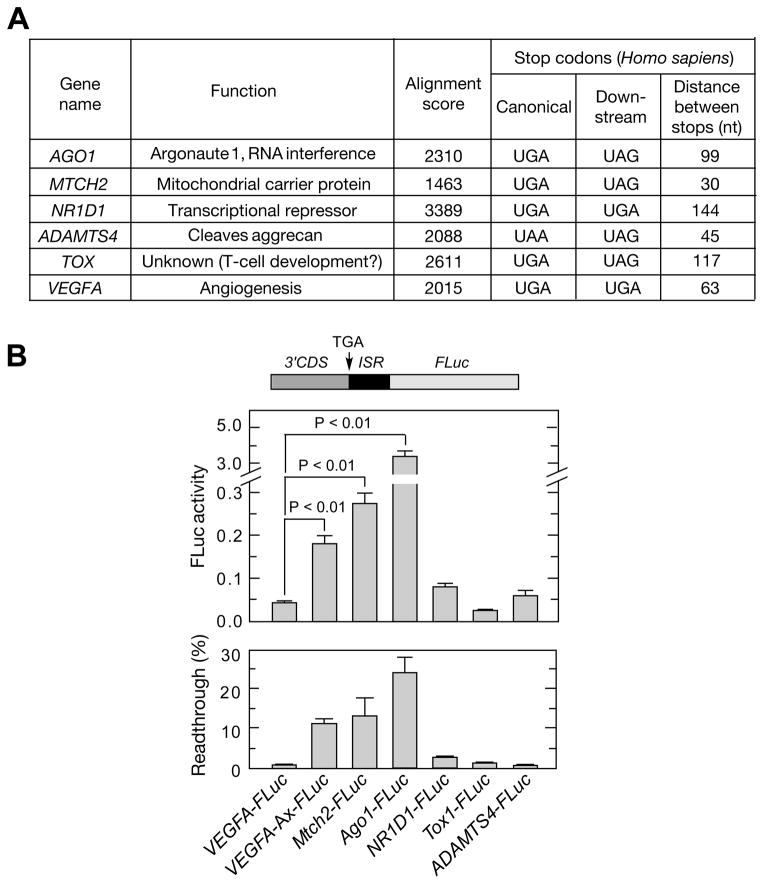
Translational readthrough, observed primarily in less complex organisms from viruses to Drosophila, expands the proteome by translating select transcripts beyond the canonical stop codon. Here, we show that vascular endothelial growth factor A (VEGFA) mRNA in mammalian endothelial cells undergoes programmed translational readthrough (PTR) generating VEGF-Ax, an isoform containing a unique 22-amino-acid C terminus extension. A cis-acting element in the VEGFA 3' UTR serves a dual function, not only encoding the appended peptide but also directing the PTR by decoding the UGA stop codon as serine. Heterogeneous nuclear ribonucleoprotein (hnRNP) A2/B1 binds this element and promotes readthrough. Remarkably, VEGF-Ax exhibits antiangiogenic activity in contrast to the proangiogenic activity of VEGF-A. Pathophysiological significance of VEGF-Ax is indicated by robust expression in multiple human tissues but depletion in colon adenocarcinoma. Furthermore, genome-wide analysis revealed AGO1 and MTCH2 as authentic readthrough targets. Overall, our studies reveal a novel protein-regulated PTR event in a vertebrate system.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
None of the other authors have any financial conflict of interest with the information in this manuscript.
Figures
Comment in
-
Cancer genetics: Ignoring the signs.Nat Rev Cancer. 2014 Aug;14(8):514. doi: 10.1038/nrc3787. Epub 2014 Jul 10. Nat Rev Cancer. 2014. PMID: 25008265 No abstract available.
References
-
- Ausprunk DH, Folkman J. Migration and proliferation of endothelial cells in preformed and newly formed blood vessels during tumor angiogenesis. Microvasc Res. 1977;14:53–65. - PubMed
-
- Bates DO, Cui TG, Doughty JM, Winkler M, Sugiono M, Shields JD, Peat D, Gillatt D, Harper SJ. VEGF165b, an inhibitory splice variant of vascular endothelial growth factor, is down-regulated in renal cell carcinoma. Cancer Res. 2002;62:4123–4131. - PubMed
-
- Carmeliet P, Ferreira V, Breier G, Pollefeyt S, Kieckens L, Gertsenstein M, Fahrig M, Vandenhoeck A, Harpal K, Eberhardt C, et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature. 1996;380:435–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
