

Structure of a RING E3 trapped in action reveals ligation mechanism for the ubiquitin-like protein NEDD8
- PMID: 24949976
- PMCID: PMC4247792
- DOI: 10.1016/j.cell.2014.04.037
Structure of a RING E3 trapped in action reveals ligation mechanism for the ubiquitin-like protein NEDD8
Abstract
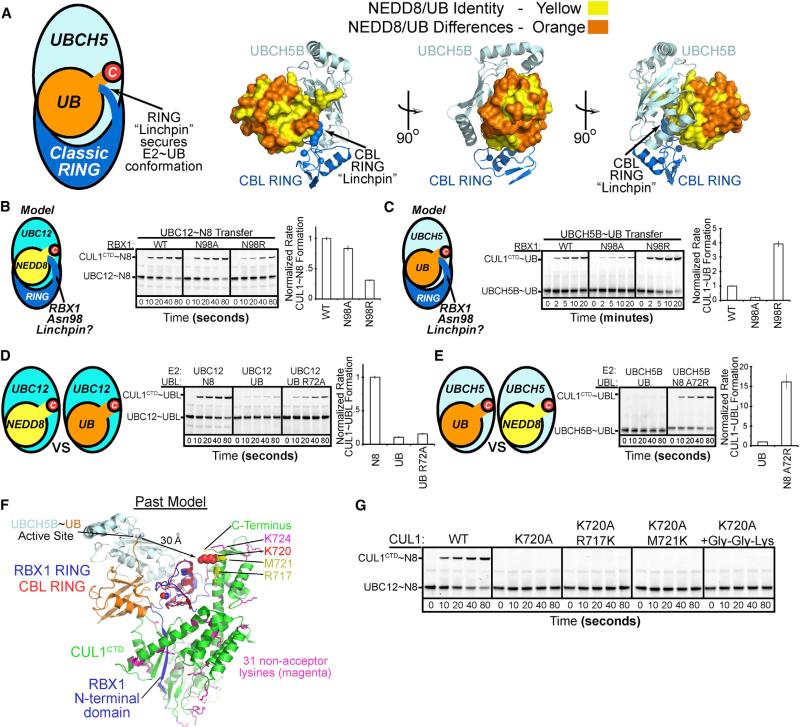
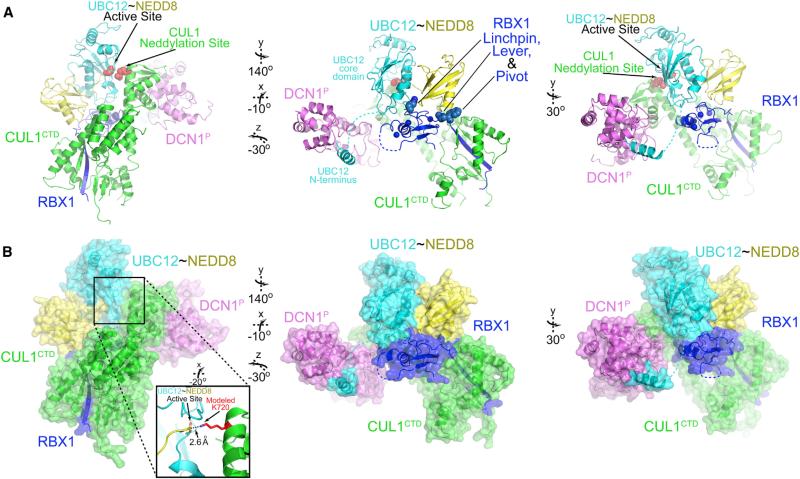
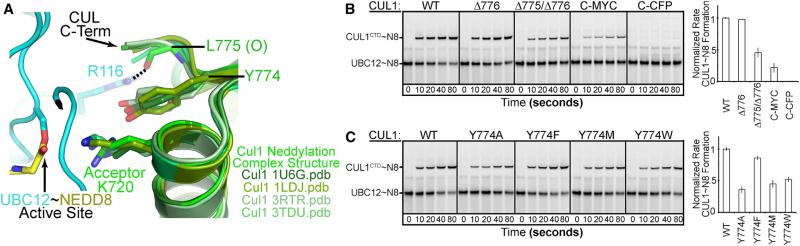
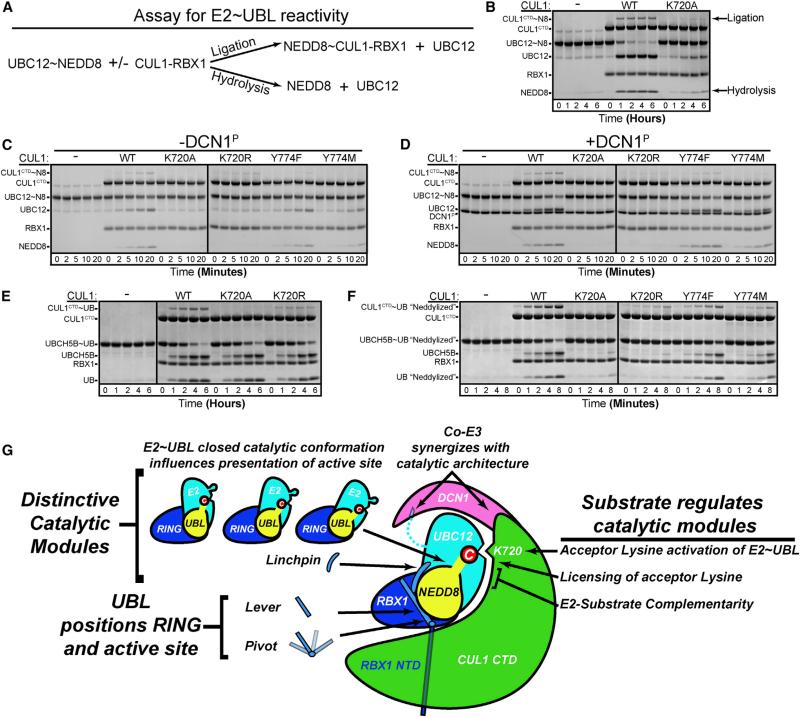
Most E3 ligases use a RING domain to activate a thioester-linked E2∼ubiquitin-like protein (UBL) intermediate and promote UBL transfer to a remotely bound target protein. Nonetheless, RING E3 mechanisms matching a specific UBL and acceptor lysine remain elusive, including for RBX1, which mediates NEDD8 ligation to cullins and >10% of all ubiquitination. We report the structure of a trapped RING E3-E2∼UBL-target intermediate representing RBX1-UBC12∼NEDD8-CUL1-DCN1, which reveals the mechanism of NEDD8 ligation and how a particular UBL and acceptor lysine are matched by a multifunctional RING E3. Numerous mechanisms specify cullin neddylation while preventing noncognate ubiquitin ligation. Notably, E2-E3-target and RING-E2∼UBL modules are not optimized to function independently, but instead require integration by the UBL and target for maximal reactivity. The UBL and target regulate the catalytic machinery by positioning the RING-E2∼UBL catalytic center, licensing the acceptor lysine, and influencing E2 reactivity, thereby driving their specific coupling by a multifunctional RING E3.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures



Similar articles
-
A RING E3-substrate complex poised for ubiquitin-like protein transfer: structural insights into cullin-RING ligases.Nat Struct Mol Biol. 2011 Jul 17;18(8):947-9. doi: 10.1038/nsmb.2086. Nat Struct Mol Biol. 2011. PMID: 21765416 Free PMC article.
-
Identification of conjugation specificity determinants unmasks vestigial preference for ubiquitin within the NEDD8 E2.Nat Struct Mol Biol. 2008 Mar;15(3):280-7. doi: 10.1038/nsmb.1387. Epub 2008 Feb 10. Nat Struct Mol Biol. 2008. PMID: 18264111
-
A dual E3 mechanism for Rub1 ligation to Cdc53.Mol Cell. 2010 Sep 10;39(5):784-96. doi: 10.1016/j.molcel.2010.08.030. Mol Cell. 2010. PMID: 20832729 Free PMC article.
-
NEDD8 and ubiquitin ligation by cullin-RING E3 ligases.Curr Opin Struct Biol. 2021 Apr;67:101-109. doi: 10.1016/j.sbi.2020.10.007. Epub 2020 Nov 5. Curr Opin Struct Biol. 2021. PMID: 33160249 Free PMC article. Review.
-
Nedd8 on cullin: building an expressway to protein destruction.Oncogene. 2004 Mar 15;23(11):1985-97. doi: 10.1038/sj.onc.1207414. Oncogene. 2004. PMID: 15021886 Review.
Cited by
-
The biological functions of Naa10 - From amino-terminal acetylation to human disease.Gene. 2015 Aug 10;567(2):103-31. doi: 10.1016/j.gene.2015.04.085. Epub 2015 May 16. Gene. 2015. PMID: 25987439 Free PMC article. Review.
-
Analysis of ubiquitin recognition by the HECT ligase E6AP provides insight into its linkage specificity.J Biol Chem. 2019 Apr 12;294(15):6113-6129. doi: 10.1074/jbc.RA118.007014. Epub 2019 Feb 8. J Biol Chem. 2019. PMID: 30737286 Free PMC article.
-
DNA activates the Nse2/Mms21 SUMO E3 ligase in the Smc5/6 complex.EMBO J. 2018 Jun 15;37(12):e98306. doi: 10.15252/embj.201798306. Epub 2018 May 16. EMBO J. 2018. PMID: 29769404 Free PMC article.
-
Targeting NEDDylation as a Novel Approach to Improve the Treatment of Head and Neck Cancer.Cancers (Basel). 2021 Jun 29;13(13):3250. doi: 10.3390/cancers13133250. Cancers (Basel). 2021. PMID: 34209641 Free PMC article. Review.
-
Cullin-RING ubiquitin E3 ligase regulation by the COP9 signalosome.Nature. 2016 Mar 31;531(7596):598-603. doi: 10.1038/nature17416. Nature. 2016. PMID: 27029275
References
-
- Angers S, Li T, Yi X, MacCoss MJ, Moon RT, Zheng N. Molecular architecture and assembly of the DDB1-CUL4A ubiquitin ligase machinery. Nature. 2006;443:590–593. - PubMed
-
- Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD. Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell. 2002;108:345–356. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
