

From molecule to mind: an integrative perspective on odor intensity
- PMID: 24950600
- PMCID: PMC4119848
- DOI: 10.1016/j.tins.2014.05.005
From molecule to mind: an integrative perspective on odor intensity
Abstract
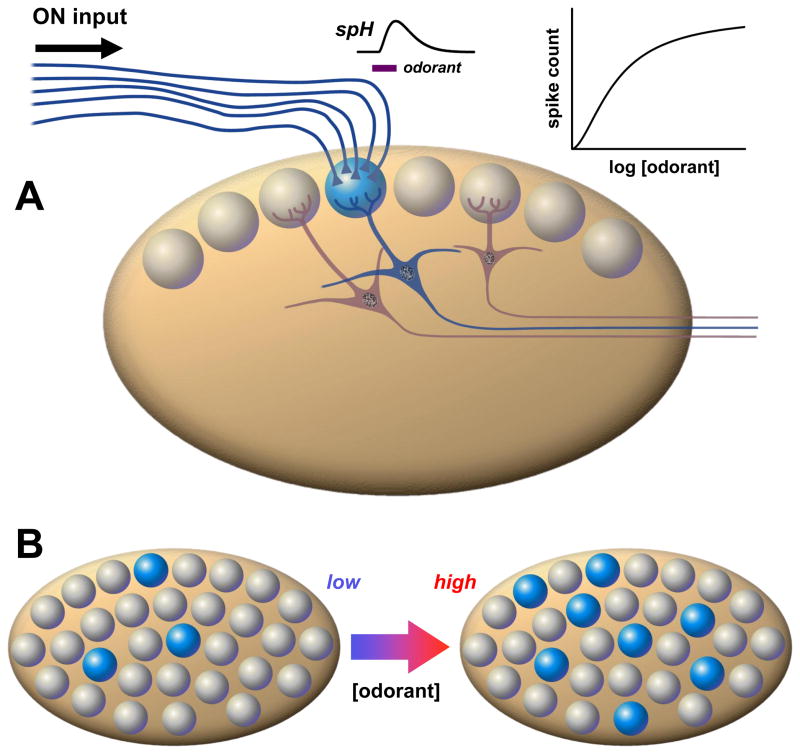
A fundamental problem in systems neuroscience is mapping the physical properties of a stimulus to perceptual characteristics. In vision, wavelength translates into color; in audition, frequency translates into pitch. Although odorant concentration is a key feature of olfactory stimuli, we do not know how concentration is translated into perceived intensity by the olfactory system. A variety of neural responses at several levels of processing have been reported to vary with odorant concentration, suggesting specific coding models. However, it remains unclear which, if any, of these phenomena underlie the perception of odor intensity. Here, we provide an overview of current models at different stages of olfactory processing, and identify promising avenues for future research.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
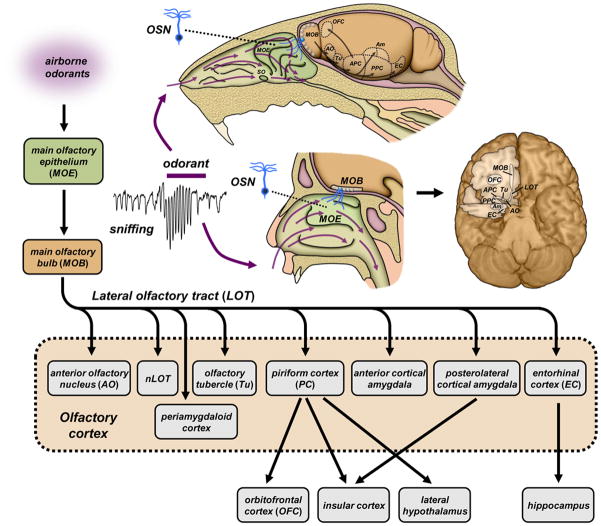
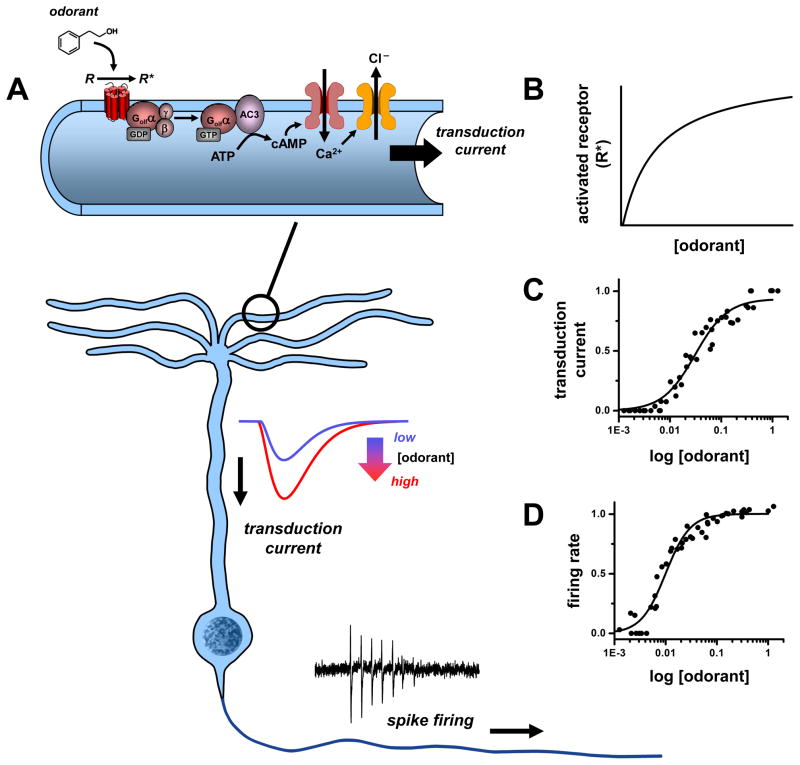
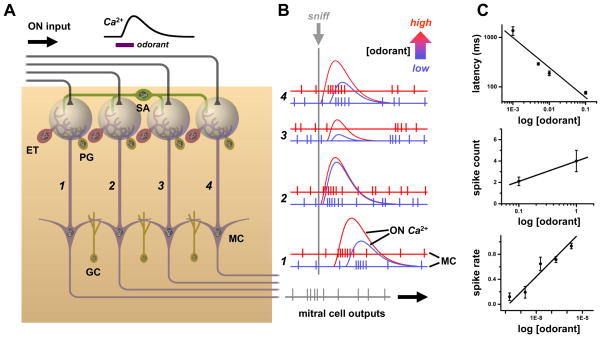
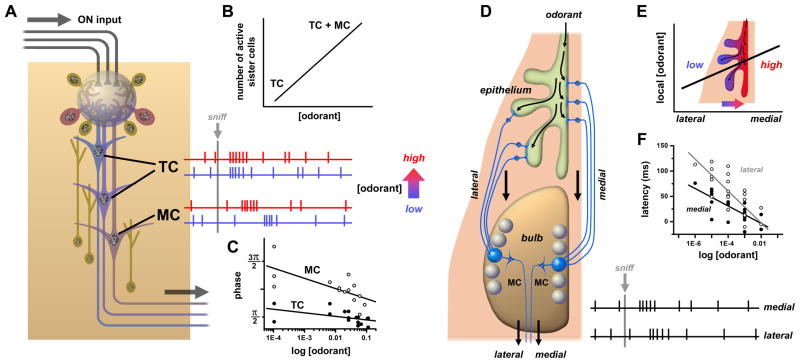
References
-
- Chastrette M, et al. Modelling the human olfactory stimulus-response function. Chem Senses. 1998;23:181–196. - PubMed
-
- Henion KE. Odor pleasantness and intensity: a single dimension? J Exp Psychol. 1971;90:275–279. - PubMed
-
- Gross-Isseroff R, Lancet D. Concentration-dependent changes of perceived odor quality. Chem Senses. 1988;13:191.
-
- Wojcik PT, Sirotin YB. Single scale for odor intensity in rat olfaction. Current biology : CB. 2014;24:568–573. - PubMed
-
- Eichenbaum H, et al. Selective olfactory deficits in case H.M. Brain. 1983;106 (Pt 2):459–472. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
