

AMPK is involved in mediation of erythropoietin influence on metabolic activity and reactive oxygen species production in white adipocytes
- PMID: 24953559
- PMCID: PMC4160370
- DOI: 10.1016/j.biocel.2014.06.008
AMPK is involved in mediation of erythropoietin influence on metabolic activity and reactive oxygen species production in white adipocytes
Abstract
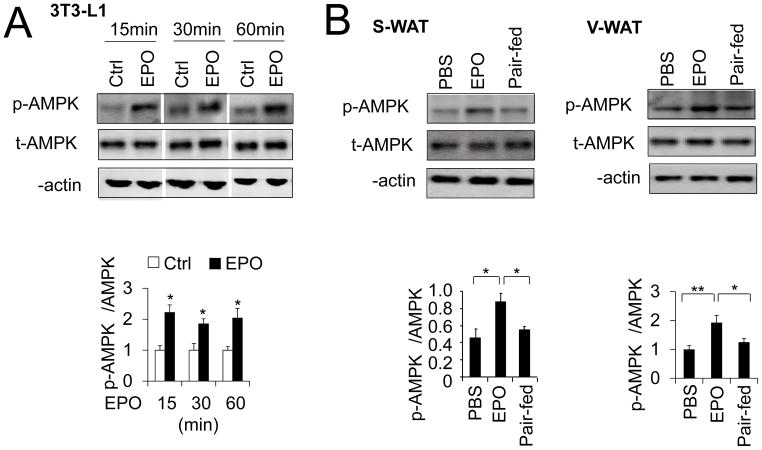
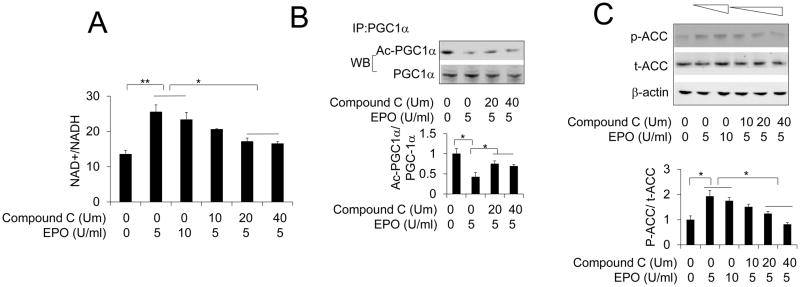
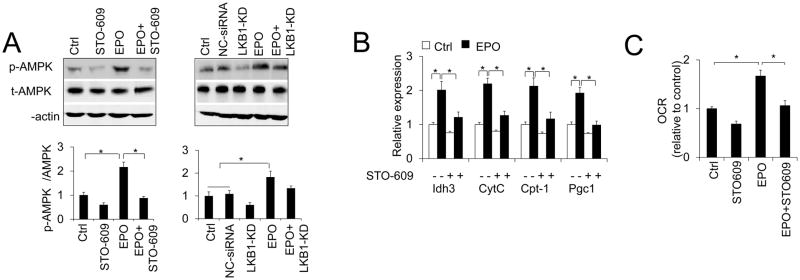
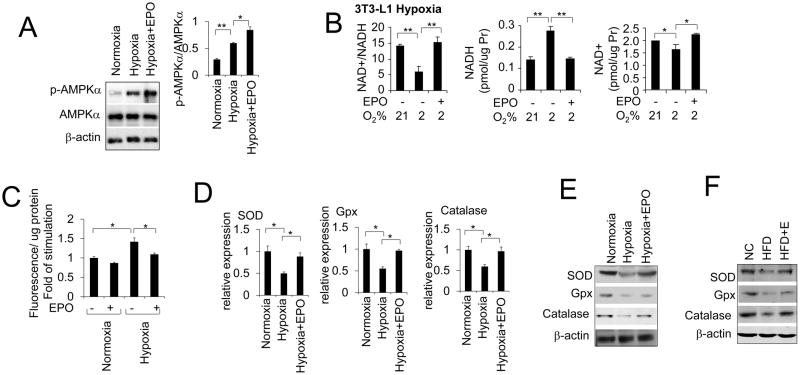
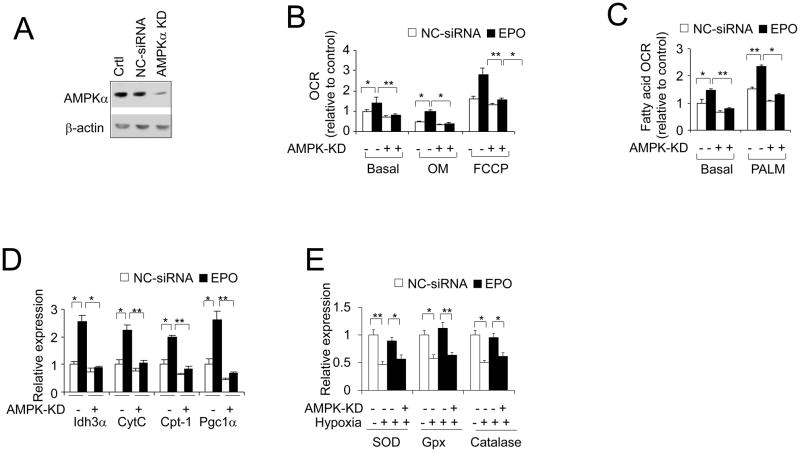
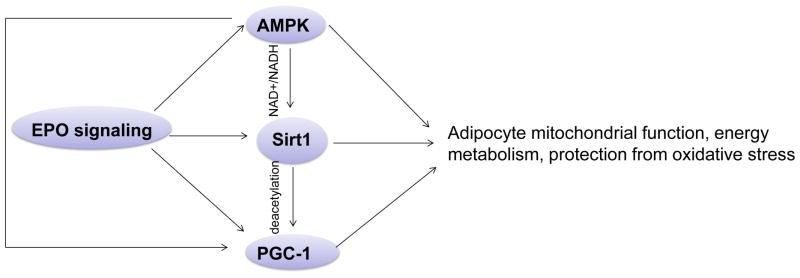
Erythropoietin, discovered for its indispensable role during erythropoiesis, has been used in therapy for selected red blood cell disorders in erythropoietin-deficient patients. The biological activities of erythropoietin have been found in animal models to extend to non-erythroid tissues due to the expression of erythropoietin receptor. We previously demonstrated that erythropoietin promotes metabolic activity and white adipocytes browning to increase mitochondrial function and energy expenditure via peroxisome proliferator-activated receptor alpha and Sirtuin1. Here we report that AMP-activated protein kinase was activated by erythropoietin possibly via Ca(2+)/calmodulin-dependent protein kinase kinase in adipocytes as well as in white adipose tissue from diet induced obese mice. Erythropoietin increased cellular nicotinamide adenine dinucleotide via increased AMP-activated protein kinase activity, possibly leading to Sirtuin1 activation. AMP-activated protein kinase knock down reduced erythropoietin mediated increase in cellular oxidative function including the increased oxygen consumption rate, fatty acid utilization and induction of key metabolic genes. Under hypoxia, adipocytes were found to generate more reactive oxygen species, and erythropoietin reduced the reactive oxygen species and increased antioxidant gene expression, suggesting that erythropoietin may provide protection from oxidative stress in adipocytes. Erythropoietin also reversed increased nicotinamide adenine dinucleotide by hypoxia via increased AMP-activated protein kinase. Additionally, AMP-activated protein kinase is found to be involved in erythropoietin stimulated increase in oxygen consumption rate, fatty acid oxidation and mitochondrial gene expression. AMP-activated protein kinase knock down impaired erythropoietin stimulated increases in antioxidant gene expression. Collectively, our findings identify the AMP-activated protein kinase involvement in erythropoietin signaling in regulating adipocyte cellular redox status and metabolic activity.
Keywords: AMP-activated protein kinase; Adipocytes; Erythropoietin; Oxidative metabolism; Reactive oxygen species.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
Similar articles
-
The diabetes medication canagliflozin promotes mitochondrial remodelling of adipocyte via the AMPK-Sirt1-Pgc-1α signalling pathway.Adipocyte. 2020 Dec;9(1):484-494. doi: 10.1080/21623945.2020.1807850. Adipocyte. 2020. PMID: 32835596 Free PMC article.
-
Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK-SIRT1-PGC-1alpha pathway.Proc Natl Acad Sci U S A. 2010 Jul 13;107(28):12553-8. doi: 10.1073/pnas.1006962107. Epub 2010 Jun 28. Proc Natl Acad Sci U S A. 2010. PMID: 20616029 Free PMC article.
-
4-hydroxybenzoic acid induces browning of white adipose tissue through the AMPK-DRP1 pathway in HFD-induced obese mice.Phytomedicine. 2025 Feb;137:156353. doi: 10.1016/j.phymed.2024.156353. Epub 2024 Dec 27. Phytomedicine. 2025. PMID: 39799892
-
Regulatory roles for L-arginine in reducing white adipose tissue.Front Biosci (Landmark Ed). 2012 Jun 1;17(6):2237-46. doi: 10.2741/4047. Front Biosci (Landmark Ed). 2012. PMID: 22652774 Free PMC article. Review.
-
Perivascular fat, AMP-activated protein kinase and vascular diseases.Br J Pharmacol. 2014 Feb;171(3):595-617. doi: 10.1111/bph.12479. Br J Pharmacol. 2014. PMID: 24490856 Free PMC article. Review.
Cited by
-
The impact of aging and oxidative stress in metabolic and nervous system disorders: programmed cell death and molecular signal transduction crosstalk.Front Immunol. 2023 Nov 8;14:1273570. doi: 10.3389/fimmu.2023.1273570. eCollection 2023. Front Immunol. 2023. PMID: 38022638 Free PMC article. Review.
-
Targeting molecules to medicine with mTOR, autophagy and neurodegenerative disorders.Br J Clin Pharmacol. 2016 Nov;82(5):1245-1266. doi: 10.1111/bcp.12804. Epub 2015 Dec 26. Br J Clin Pharmacol. 2016. PMID: 26469771 Free PMC article. Review.
-
Innovative therapeutic strategies for cardiovascular disease.EXCLI J. 2023 Jul 26;22:690-715. doi: 10.17179/excli2023-6306. eCollection 2023. EXCLI J. 2023. PMID: 37593239 Free PMC article. Review.
-
Novel nervous and multi-system regenerative therapeutic strategies for diabetes mellitus with mTOR.Neural Regen Res. 2016 Mar;11(3):372-85. doi: 10.4103/1673-5374.179032. Neural Regen Res. 2016. PMID: 27127460 Free PMC article. Review.
-
Regeneration in the nervous system with erythropoietin.Front Biosci (Landmark Ed). 2016 Jan;21(3):561-596. doi: 10.2741/4408. Front Biosci (Landmark Ed). 2016. PMID: 26549969 Free PMC article.
References
-
- Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414:813–820. - PubMed
-
- Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clinical chemistry. 2009;55:611–622. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
