

Cold hypersensitivity increases with age in mice with sickle cell disease
- PMID: 24953902
- PMCID: PMC4250326
- DOI: 10.1016/j.pain.2014.05.030
Cold hypersensitivity increases with age in mice with sickle cell disease
Abstract
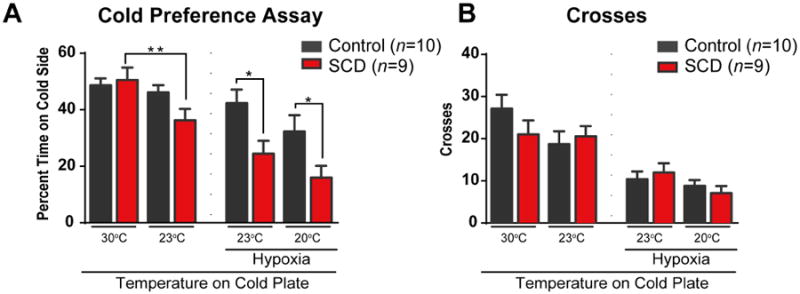
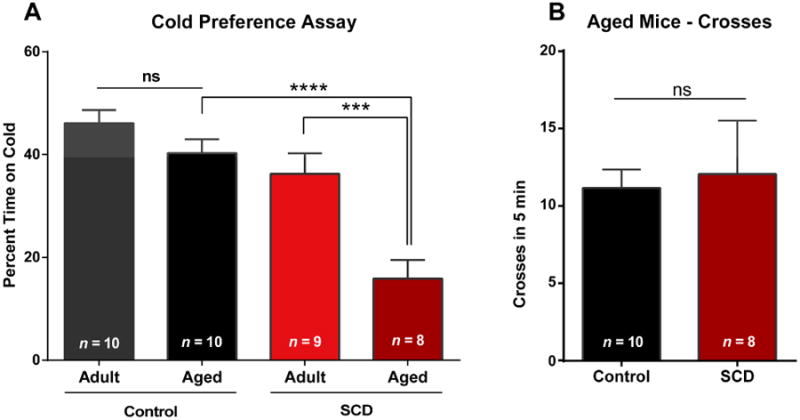
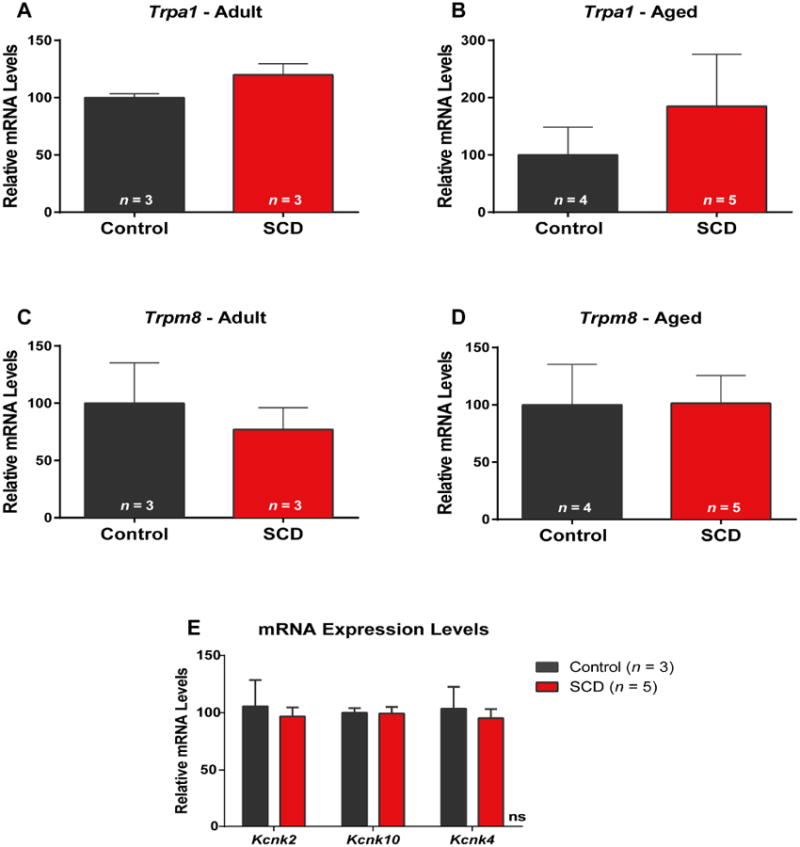
Sickle cell disease (SCD) is associated with acute vaso-occlusive crises that trigger painful episodes and frequently involves ongoing, chronic pain. In addition, both humans and mice with SCD experience heightened cold sensitivity. However, studies have not addressed the mechanism(s) underlying the cold sensitization or its progression with age. Here we measured thermotaxis behavior in young and aged mice with severe SCD. Sickle mice had a marked increase in cold sensitivity measured by a cold preference test. Furthermore, cold hypersensitivity worsened with advanced age. We assessed whether enhanced peripheral input contributes to the chronic cold pain behavior by recording from C fibers, many of which are cold sensitive, in skin-nerve preparations. We observed that C fibers from sickle mice displayed a shift to warmer (more sensitive) cold detection thresholds. To address mechanisms underlying the cold sensitization in primary afferent neurons, we quantified mRNA expression levels for ion channels thought to be involved in cold detection. These included the transient receptor potential melastatin 8 (Trpm8) and transient receptor potential ankyrin 1 (Trpa1) channels, as well as the 2-pore domain potassium channels, TREK-1 (Kcnk2), TREK-2 (Kcnk10), and TRAAK (Kcnk4). Surprisingly, transcript expression levels of all of these channels were comparable between sickle and control mice. We further examined transcript expression of 83 additional pain-related genes, and found increased mRNA levels for endothelin 1 and tachykinin receptor 1. These factors may contribute to hypersensitivity in sickle mice at both the afferent and behavioral levels.
Keywords: Aging; C fiber; Endothelin 1; KCNK; TRPA1; TRPM8.
Copyright © 2014 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Comment in
-
Does cold hypersensitivity increase with age in sickle cell disease?Pain. 2014 Dec;155(12):2439-2440. doi: 10.1016/j.pain.2014.08.010. Epub 2014 Aug 27. Pain. 2014. PMID: 25172823 Free PMC article. No abstract available.
Similar articles
-
Does cold hypersensitivity increase with age in sickle cell disease?Pain. 2014 Dec;155(12):2439-2440. doi: 10.1016/j.pain.2014.08.010. Epub 2014 Aug 27. Pain. 2014. PMID: 25172823 Free PMC article. No abstract available.
-
The preventive effect of resiniferatoxin on the development of cold hypersensitivity induced by spinal nerve ligation: involvement of TRPM8.BMC Neurosci. 2016 Jun 21;17(1):38. doi: 10.1186/s12868-016-0273-8. BMC Neurosci. 2016. PMID: 27329106 Free PMC article.
-
The contribution of TRPM8 and TRPA1 channels to cold allodynia and neuropathic pain.PLoS One. 2009 Oct 8;4(10):e7383. doi: 10.1371/journal.pone.0007383. PLoS One. 2009. PMID: 19812688 Free PMC article.
-
IKD Current in Cold Transduction and Damage-Triggered Cold Hypersensitivity.Adv Exp Med Biol. 2017;1015:265-277. doi: 10.1007/978-3-319-62817-2_14. Adv Exp Med Biol. 2017. PMID: 29080031 Review.
-
Converting cold into pain.Exp Brain Res. 2009 Jun;196(1):13-30. doi: 10.1007/s00221-009-1797-2. Epub 2009 Apr 28. Exp Brain Res. 2009. PMID: 19399484 Review.
Cited by
-
Mechanisms of pain in sickle cell disease.Br J Pain. 2021 May;15(2):213-220. doi: 10.1177/2049463720920682. Epub 2020 May 22. Br J Pain. 2021. PMID: 34055342 Free PMC article.
-
Chemokine (c-c motif) receptor 2 mediates mechanical and cold hypersensitivity in sickle cell disease mice.Pain. 2018 Aug;159(8):1652-1663. doi: 10.1097/j.pain.0000000000001253. Pain. 2018. PMID: 29697532 Free PMC article.
-
Does cold hypersensitivity increase with age in sickle cell disease?Pain. 2014 Dec;155(12):2439-2440. doi: 10.1016/j.pain.2014.08.010. Epub 2014 Aug 27. Pain. 2014. PMID: 25172823 Free PMC article. No abstract available.
-
Quantification of pain in sickle mice using facial expressions and body measurements.Blood Cells Mol Dis. 2016 Mar;57:58-66. doi: 10.1016/j.bcmd.2015.12.006. Epub 2015 Dec 14. Blood Cells Mol Dis. 2016. PMID: 26852657 Free PMC article.
-
Children and adolescents with sickle cell disease have worse cold and mechanical hypersensitivity during acute painful events.Pain. 2019 Feb;160(2):407-416. doi: 10.1097/j.pain.0000000000001407. Pain. 2019. PMID: 30247266 Free PMC article.
References
-
- Andoh T, Nagasawa T, Kuraishi Y. Expression of tachykinin NK1 receptor mRNA in dorsal root ganglia of the mouse. Brain Res Mol Brain Res. 1996;35(1-2):329–332. - PubMed
-
- Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron. 2004;41(6):849–857. - PubMed
-
- Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, Yamoah EN, Basbaum AI, Julius D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell. 2006;124(6):1269–1282. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
