

Hepatic oxidative stress promotes insulin-STAT-5 signaling and obesity by inactivating protein tyrosine phosphatase N2
- PMID: 24954415
- PMCID: PMC4335267
- DOI: 10.1016/j.cmet.2014.05.011
Hepatic oxidative stress promotes insulin-STAT-5 signaling and obesity by inactivating protein tyrosine phosphatase N2
Abstract
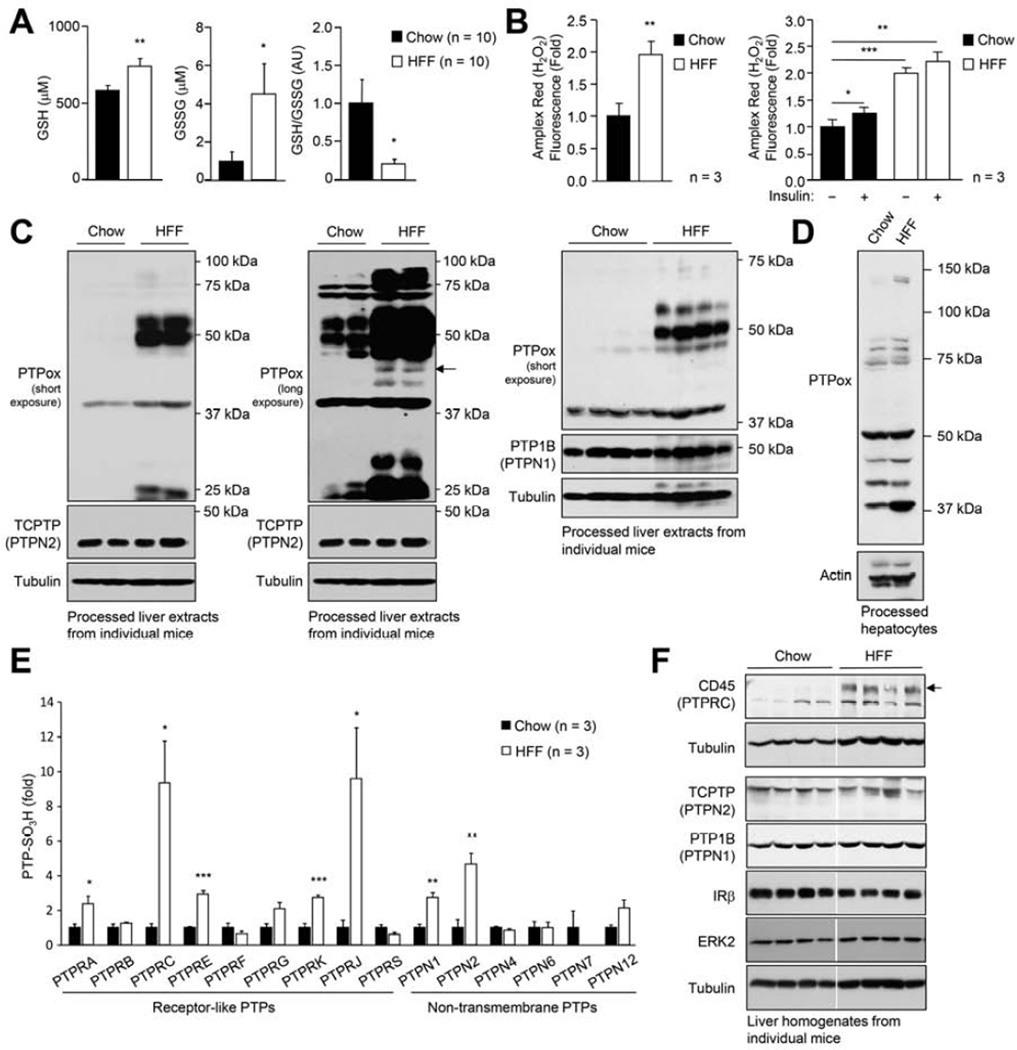
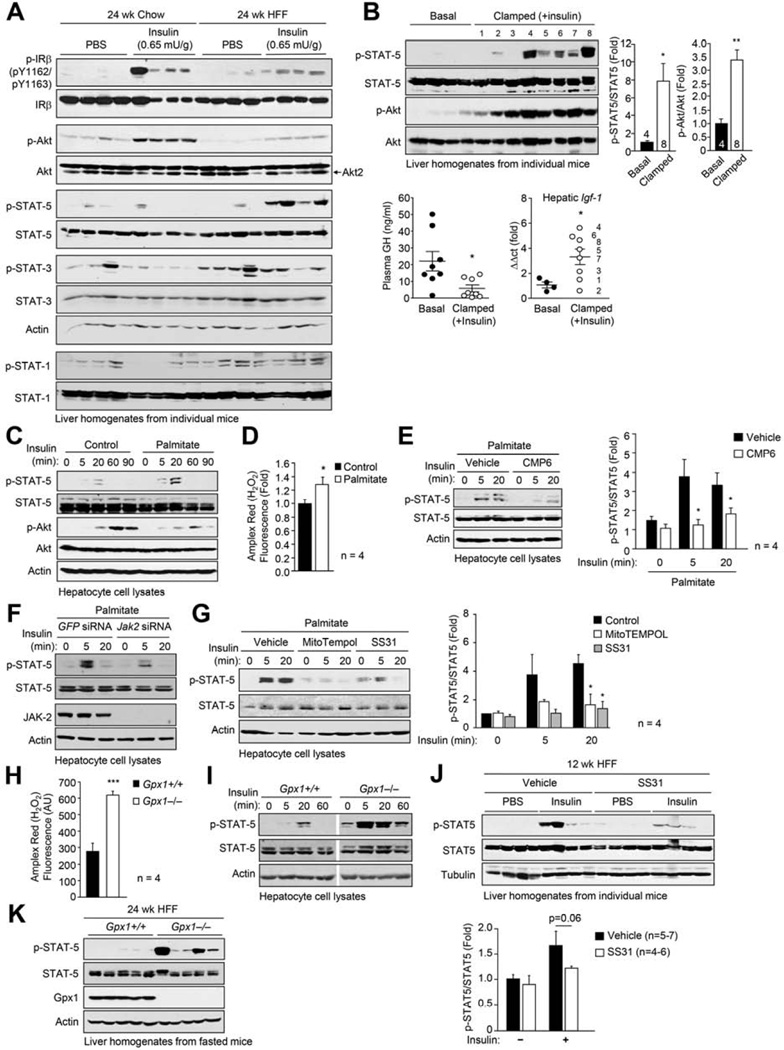
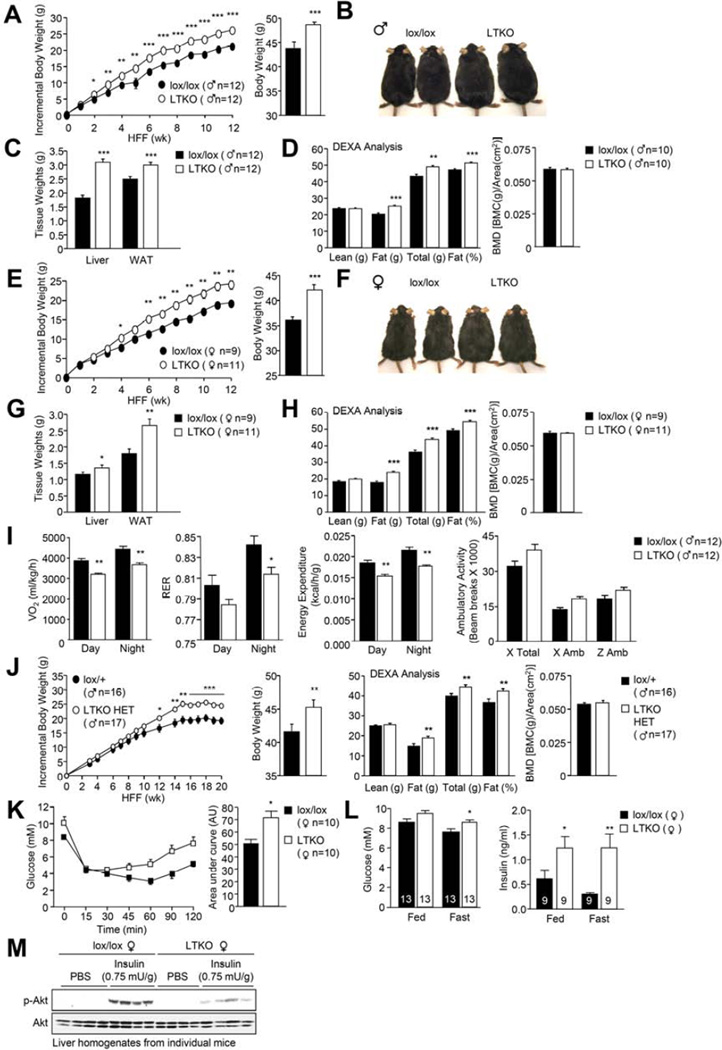
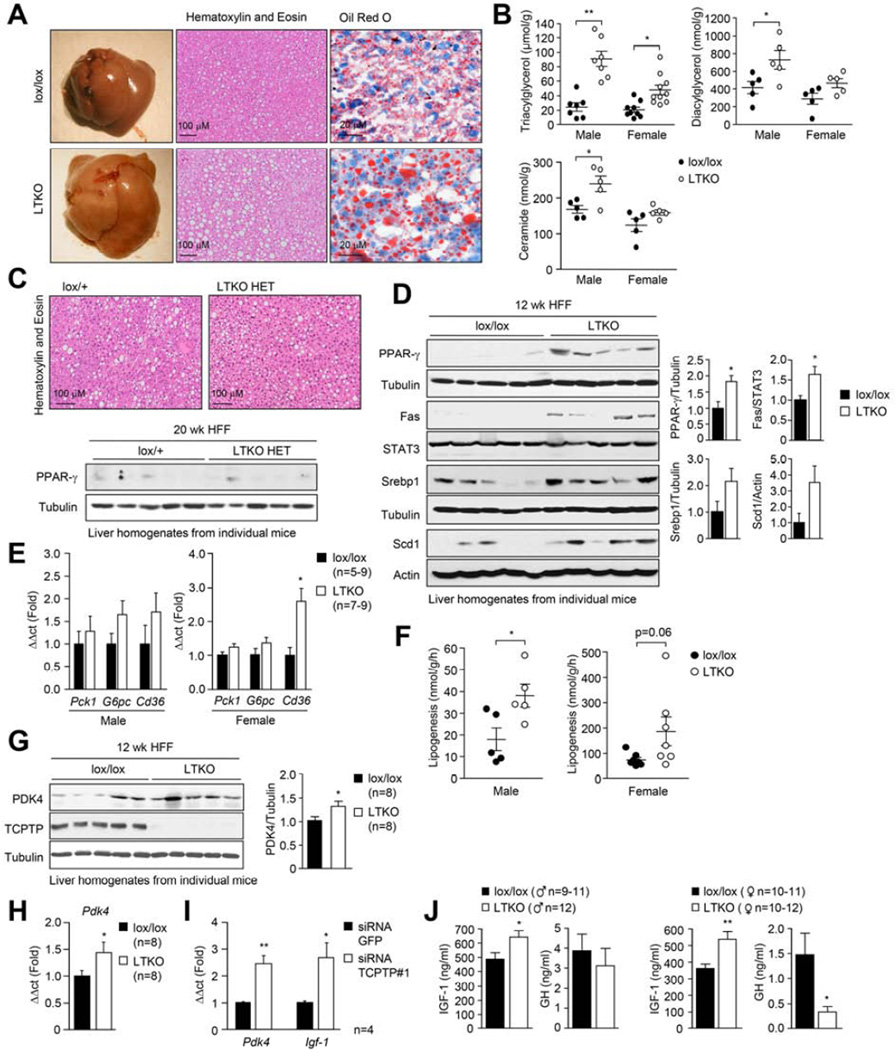
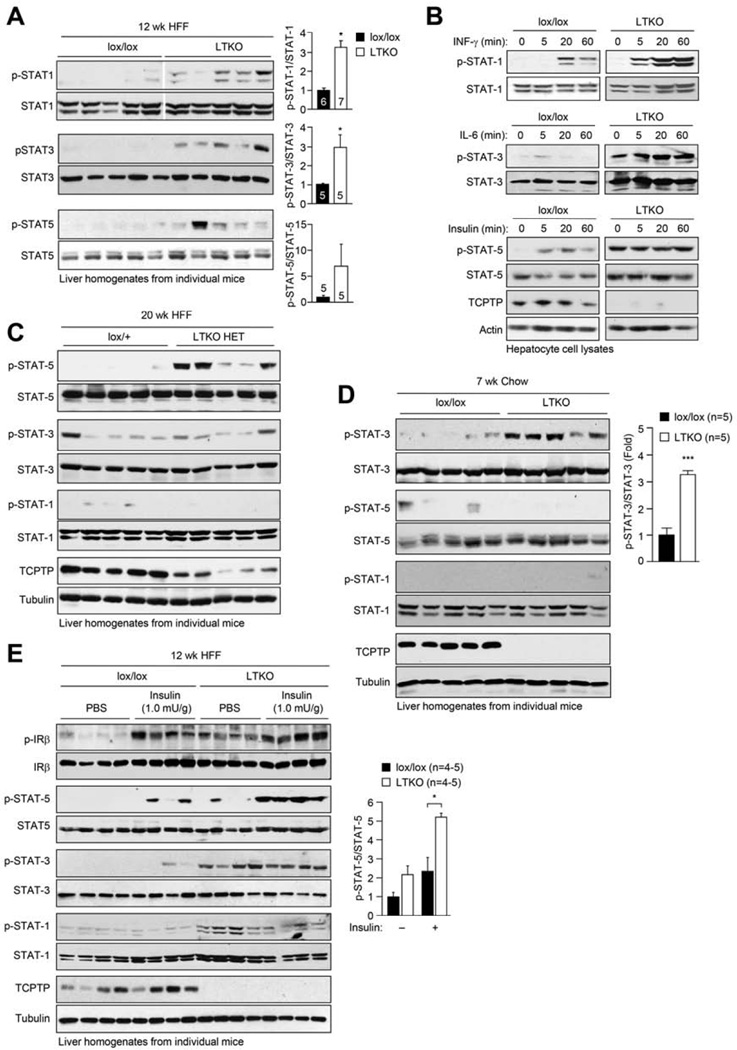
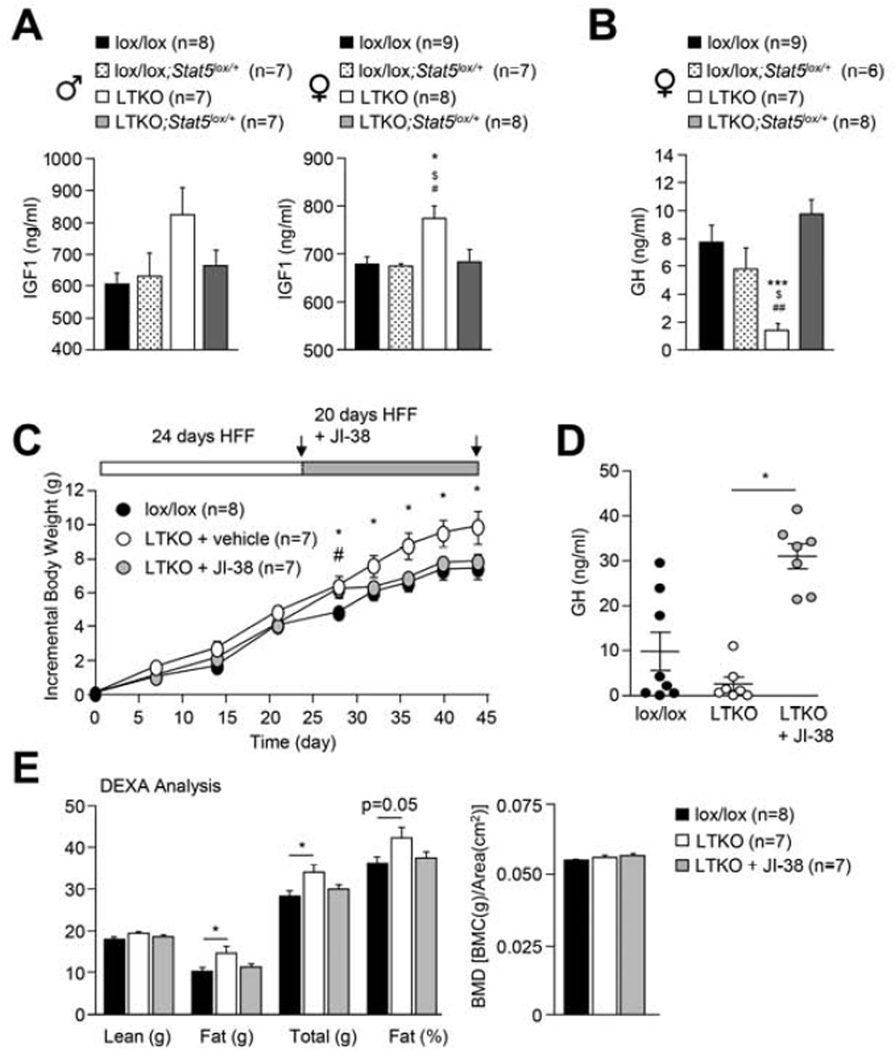
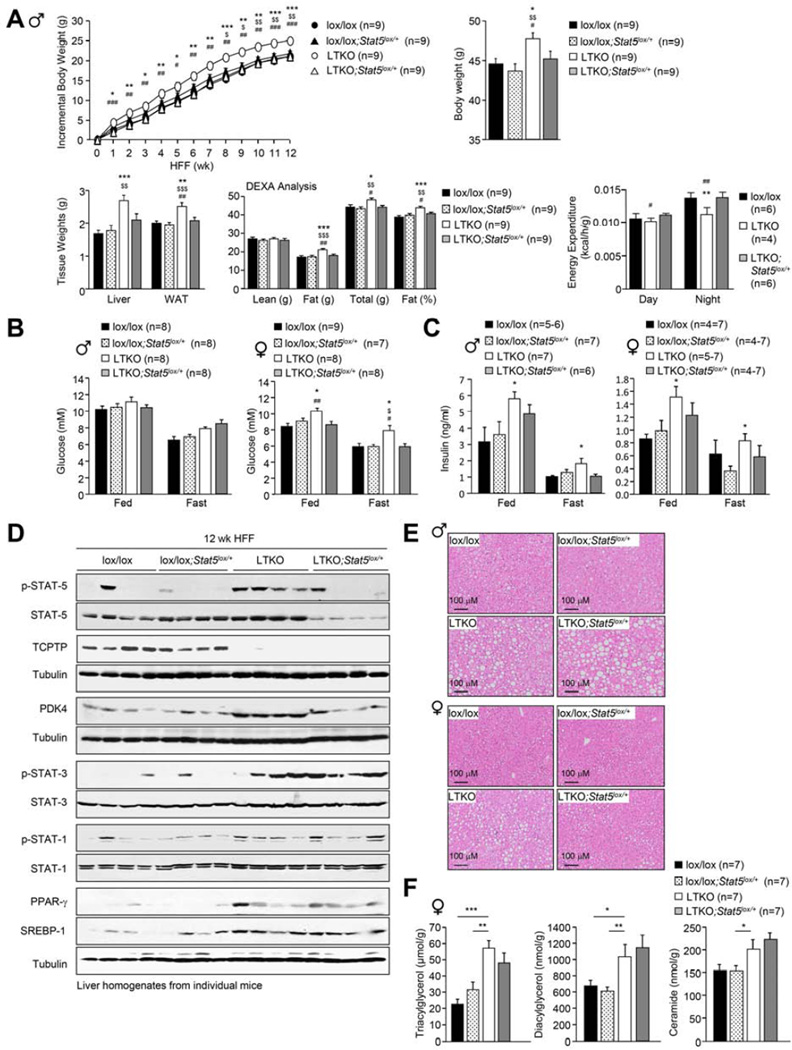
Hepatic insulin resistance is a key contributor to the pathogenesis of obesity and type 2 diabetes (T2D). Paradoxically, the development of insulin resistance in the liver is not universal, but pathway selective, such that insulin fails to suppress gluconeogenesis but promotes lipogenesis, contributing to the hyperglycemia, steatosis, and hypertriglyceridemia that underpin the deteriorating glucose control and microvascular complications in T2D. The molecular basis for the pathway-specific insulin resistance remains unknown. Here we report that oxidative stress accompanying obesity inactivates protein-tyrosine phosphatases (PTPs) in the liver to activate select signaling pathways that exacerbate disease progression. In obese mice, hepatic PTPN2 (TCPTP) inactivation promoted lipogenesis and steatosis and insulin-STAT-5 signaling. The enhanced STAT-5 signaling increased hepatic IGF-1 production, which suppressed central growth hormone release and exacerbated the development of obesity and T2D. Our studies define a mechanism for the development of selective insulin resistance with wide-ranging implications for diseases characterized by oxidative stress.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Barclay JL, Nelson CN, Ishikawa M, Murray LA, Kerr LM, McPhee TR, Powell EE, Waters MJ. GH-dependent STAT5 signaling plays an important role in hepatic lipid metabolism. Endocrinology. 2011;152:181–192. - PubMed
-
- Brown MS, Goldstein JL. Selective versus total insulin resistance: a pathogenic paradox. Cell Metab. 2008;7:95–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
