

A cell-surface β-hydroxylase is a biomarker and therapeutic target for hepatocellular carcinoma
- PMID: 24954865
- PMCID: PMC4176525
- DOI: 10.1002/hep.27275
A cell-surface β-hydroxylase is a biomarker and therapeutic target for hepatocellular carcinoma
Abstract
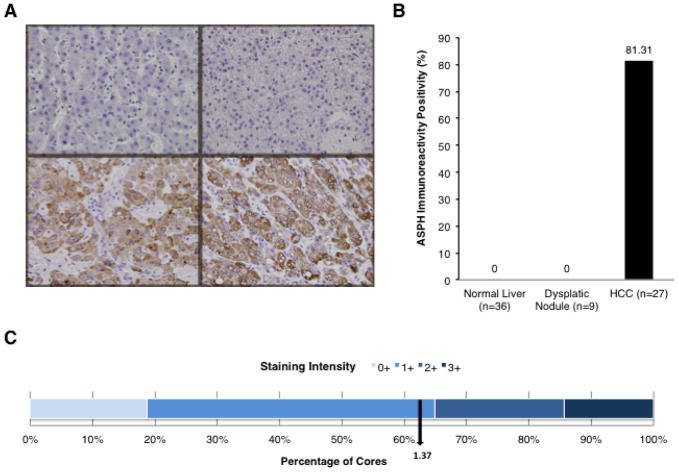
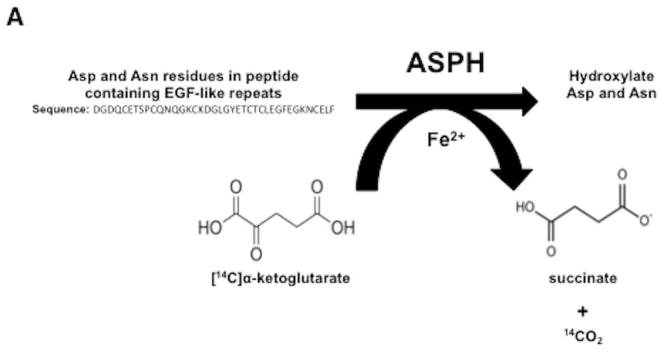
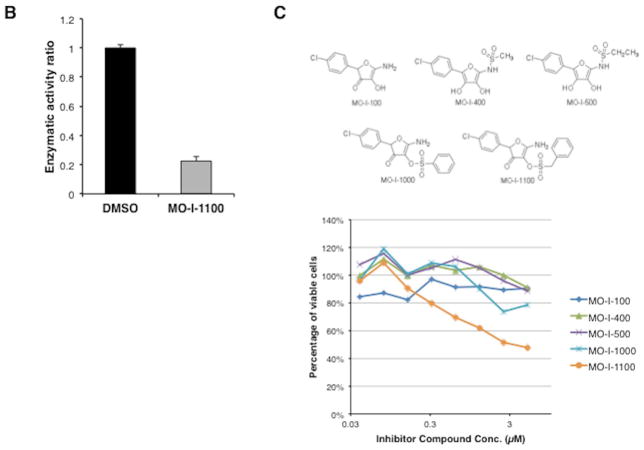
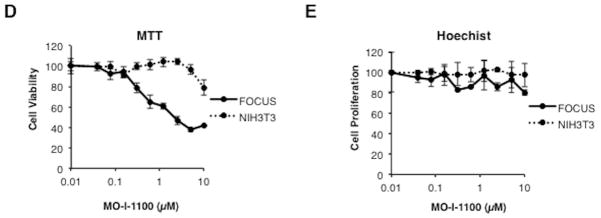
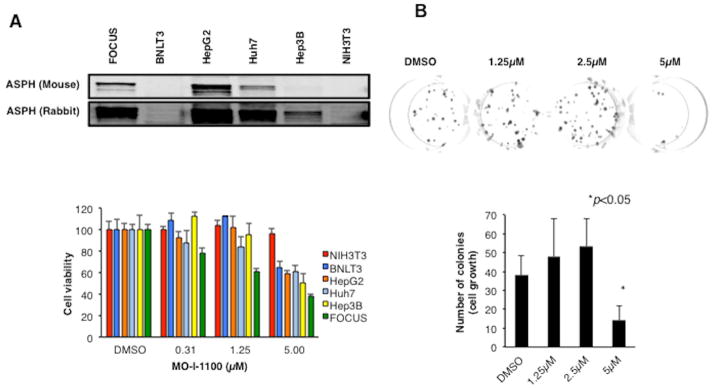
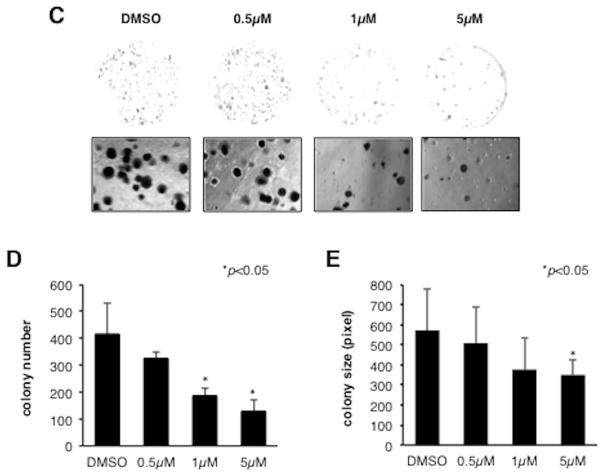
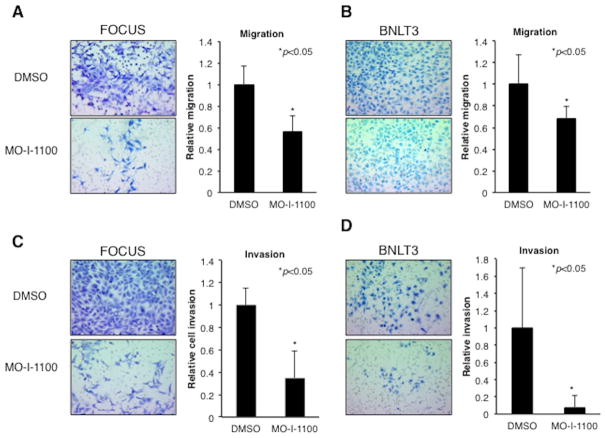
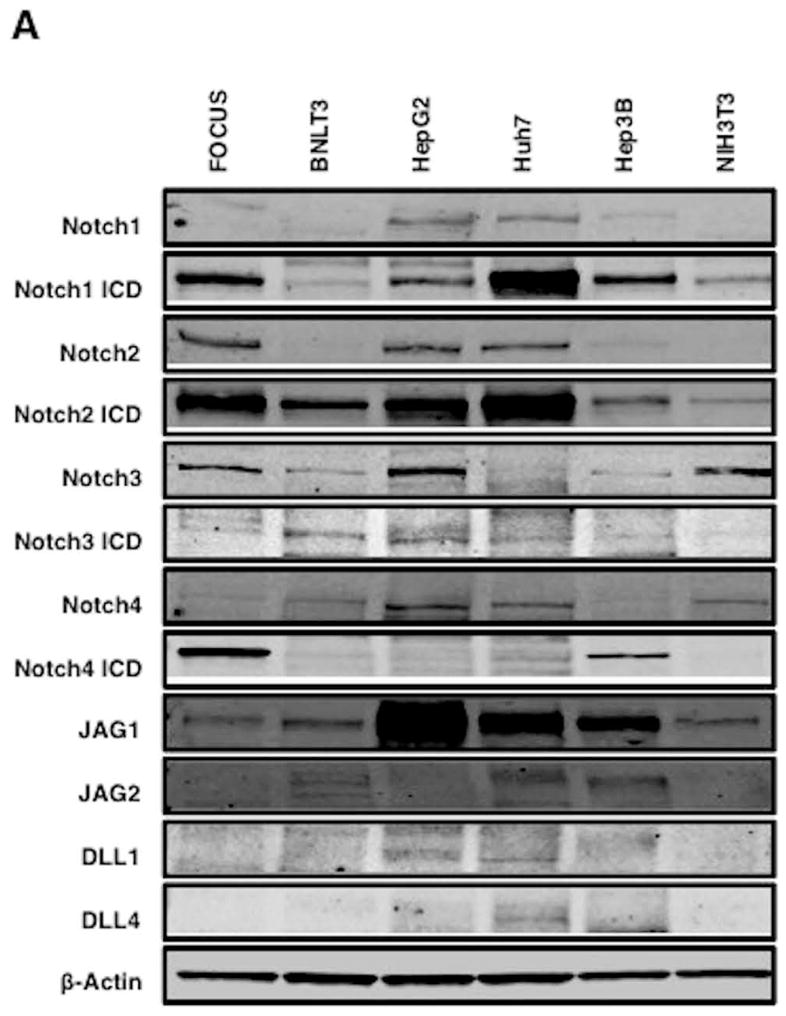
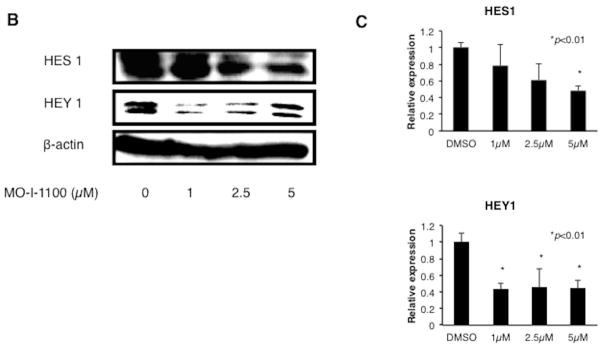
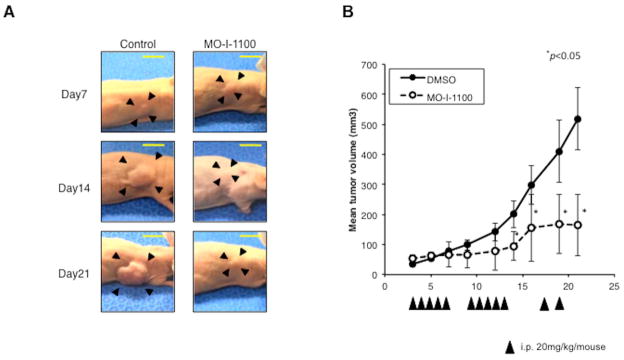
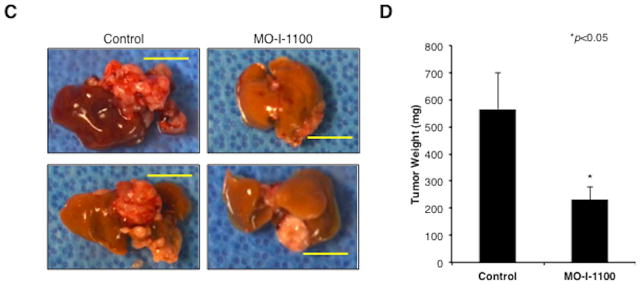
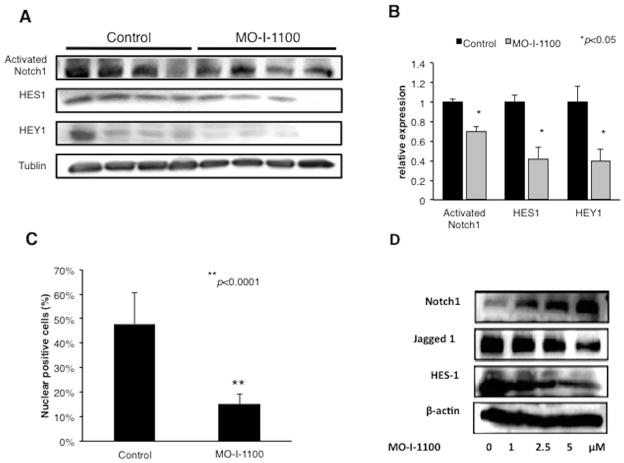
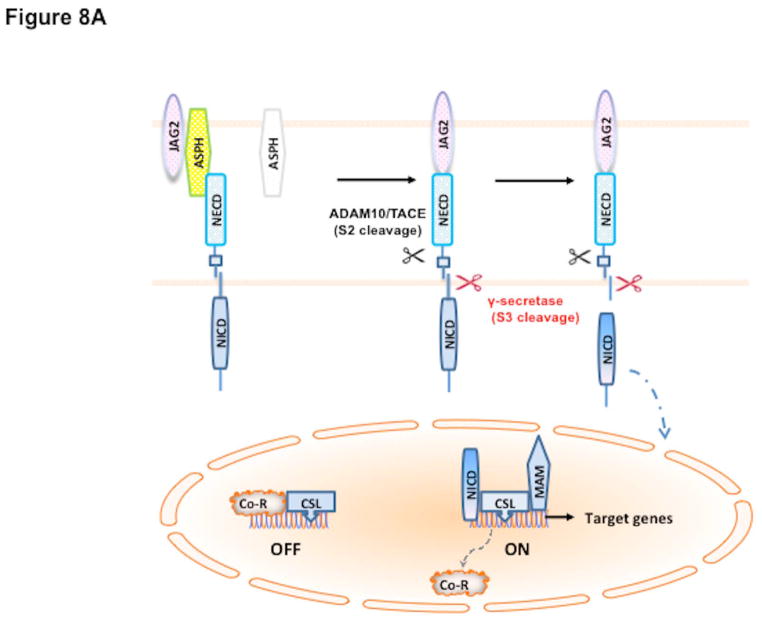
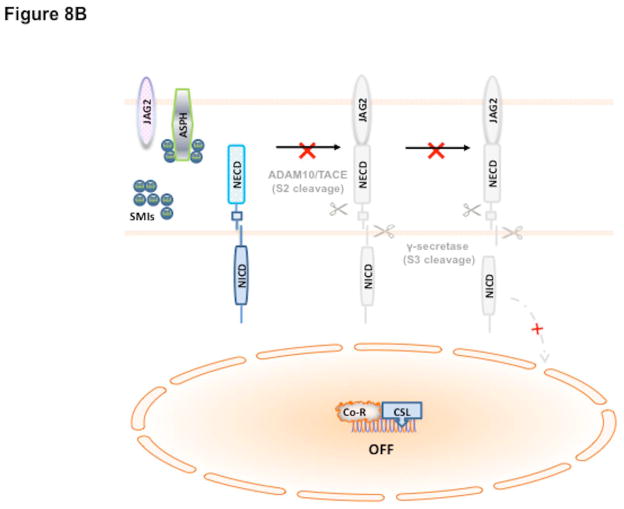
Hepatocellular carcinoma (HCC) has a poor prognosis as a result of widespread intra- and extrahepatic metastases. There is an urgent need to understand signaling cascades that promote disease progression. Aspartyl-(asparaginyl)-β-hydroxylase (ASPH) is a cell-surface enzyme that generates enhanced cell motility, migration, invasion, and metastatic spread in HCC. We hypothesize that inhibition of its enzymatic activity could have antitumor effects. Small molecule inhibitors (SMIs) were developed based on the crystal structure of the ASPH catalytic site followed by computer-assisted drug design. Candidate compounds were tested for inhibition of β-hydroxylase activity and selected for their capability to modulate cell proliferation, migration, invasion, and colony formation in vitro and to inhibit HCC tumor growth in vivo using orthotopic and subcutaneous murine models. The biological effects of SMIs on the Notch signaling cascade were evaluated. The SMI inhibitor, MO-I-1100, was selected because it reduced ASPH enzymatic activity by 80% and suppressed HCC cell migration, invasion, and anchorage-independent growth. Furthermore, substantial inhibition of HCC tumor growth and progression was observed in both animal models. The mechanism(s) for this antitumor effect was associated with reduced activation of Notch signaling both in vitro and in vivo.
Conclusions: These studies suggest that the enzymatic activity of ASPH is important for hepatic oncogenesis. Reduced β-hydroxylase activity generated by the SMI MO-I-1100 leads to antitumor effects through inhibiting Notch signaling cascade in HCC. ASPH promotes the generation of an HCC malignant phenotype and represents an attractive molecular target for therapy of this fatal disease.
© 2014 by the American Association for the Study of Liver Diseases.
Conflict of interest statement
Figures
References
-
- Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, de Oliveira AC, et al. Sorafenib in advanced hepatocellular carcinoma. N Engl J Med. 2008;359:378–390. - PubMed
-
- Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S, Kim JS, Luo R, et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: a phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009;10:25–34. - PubMed
-
- Jia S, VanDusen WJ, Diehl RE, Kohl NE, Dixon RA, Elliston KO, Stern AM, et al. cDNA cloning and expression of bovine aspartyl (asparaginyl) beta-hydroxylase. J Biol Chem. 1992;267:14322–14327. - PubMed
-
- Engel J. EGF-like domains in extracellular matrix proteins: localized signals for growth and differentiation? FEBS Lett. 1989;251:1–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical