

Direct evaluation of tRNA aminoacylation status by the T-box riboswitch using tRNA-mRNA stacking and steric readout
- PMID: 24954903
- PMCID: PMC4104367
- DOI: 10.1016/j.molcel.2014.05.017
Direct evaluation of tRNA aminoacylation status by the T-box riboswitch using tRNA-mRNA stacking and steric readout
Abstract
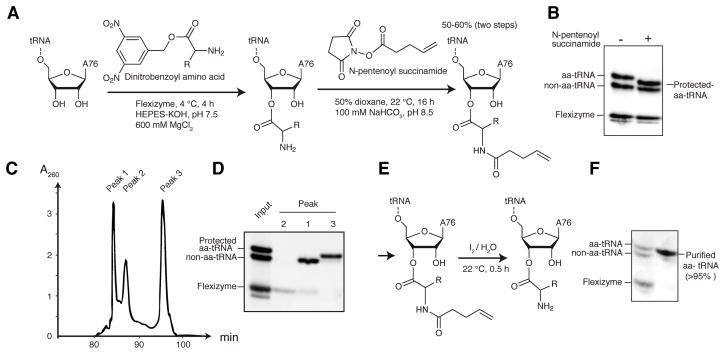
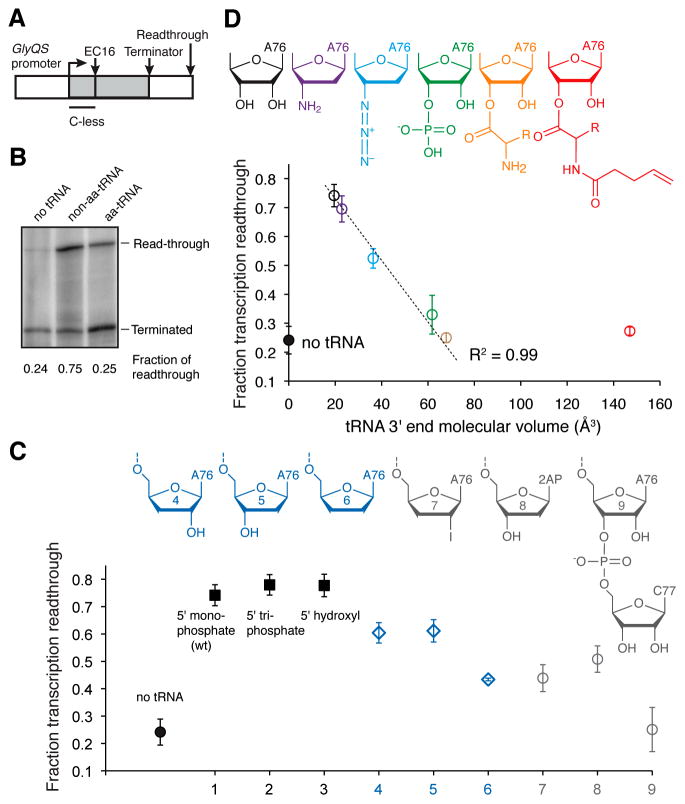
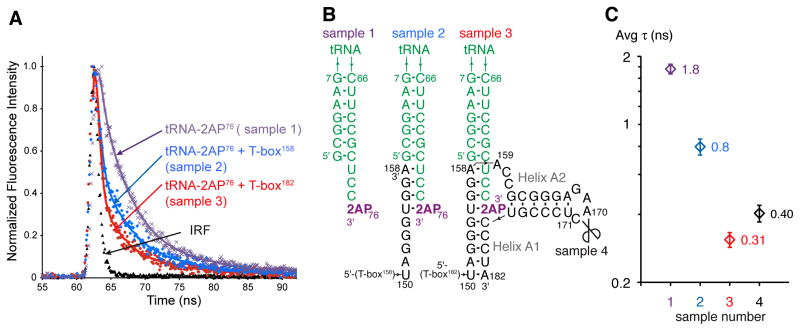
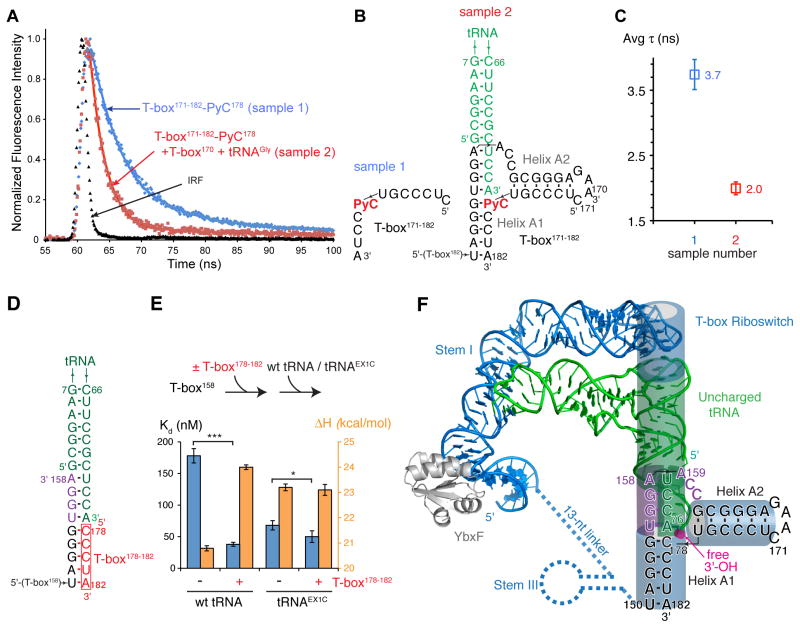
T-boxes are gene-regulatory mRNA elements with which Gram-positive bacteria sense amino acid availability. T-boxes have two functional domains. Stem I recognizes the overall shape and anticodon of tRNA, while a 3' domain evaluates its aminoacylation status, overcoming an otherwise stable transcriptional terminator if the bound tRNA is uncharged. Although T-boxes are believed to evaluate tRNA charge status without using any proteins, this has not been demonstrated experimentally because of the instability of aminoacyl-tRNA. Using a simple method to prepare homogeneous aminoacyl-tRNA, we show that the Bacillus subtilis glyQS T-box functions independently of any tRNA-binding protein. Comparison of aminoacyl-tRNA analogs demonstrates that the T-box detects the molecular volume of tRNA 3'-substituents. Calorimetry and fluorescence lifetime analysis of labeled RNAs shows that the tRNA acceptor end coaxially stacks on a helix in the T-box 3' domain. This intimate intermolecular association, selective for uncharged tRNA, stabilizes the antiterminator conformation of the T-box.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
An evolving tale of two interacting RNAs-themes and variations of the T-box riboswitch mechanism.IUBMB Life. 2019 Aug;71(8):1167-1180. doi: 10.1002/iub.2098. Epub 2019 Jun 17. IUBMB Life. 2019. PMID: 31206978 Free PMC article. Review.
-
Capture and Release of tRNA by the T-Loop Receptor in the Function of the T-Box Riboswitch.Biochemistry. 2017 Jul 18;56(28):3549-3558. doi: 10.1021/acs.biochem.7b00284. Epub 2017 Jul 3. Biochemistry. 2017. PMID: 28621923 Free PMC article.
-
Co-crystal structure of a T-box riboswitch stem I domain in complex with its cognate tRNA.Nature. 2013 Aug 15;500(7462):363-6. doi: 10.1038/nature12440. Epub 2013 Jul 28. Nature. 2013. PMID: 23892783 Free PMC article.
-
Structural basis of amino acid surveillance by higher-order tRNA-mRNA interactions.Nat Struct Mol Biol. 2019 Dec;26(12):1094-1105. doi: 10.1038/s41594-019-0326-7. Epub 2019 Nov 18. Nat Struct Mol Biol. 2019. PMID: 31740854 Free PMC article.
-
The T box mechanism: tRNA as a regulatory molecule.FEBS Lett. 2010 Jan 21;584(2):318-24. doi: 10.1016/j.febslet.2009.11.056. FEBS Lett. 2010. PMID: 19932103 Free PMC article. Review.
Cited by
-
Riboswitch-mediated regulation of riboflavin biosynthesis genes in prokaryotes.3 Biotech. 2022 Oct;12(10):278. doi: 10.1007/s13205-022-03348-3. Epub 2022 Sep 14. 3 Biotech. 2022. PMID: 36275359 Free PMC article. Review.
-
Regulation of Bacterial Gene Expression by Transcription Attenuation.Microbiol Mol Biol Rev. 2019 Jul 3;83(3):e00019-19. doi: 10.1128/MMBR.00019-19. Print 2019 Aug 21. Microbiol Mol Biol Rev. 2019. PMID: 31270135 Free PMC article. Review.
-
Hierarchical mechanism of amino acid sensing by the T-box riboswitch.Nat Commun. 2018 May 14;9(1):1896. doi: 10.1038/s41467-018-04305-6. Nat Commun. 2018. PMID: 29760498 Free PMC article.
-
New tRNA contacts facilitate ligand binding in a Mycobacterium smegmatis T box riboswitch.Proc Natl Acad Sci U S A. 2018 Apr 10;115(15):3894-3899. doi: 10.1073/pnas.1721254115. Epub 2018 Mar 26. Proc Natl Acad Sci U S A. 2018. PMID: 29581302 Free PMC article.
-
Cotranscriptional Assembly and Native Purification of Large RNA-RNA Complexes for Structural Analyses.Methods Mol Biol. 2023;2568:1-12. doi: 10.1007/978-1-0716-2687-0_1. Methods Mol Biol. 2023. PMID: 36227558 Free PMC article.
References
-
- Buttner K, Bernhardt J, Scharf C, Schmid R, Mader U, Eymann C, Antelmann H, Volker A, Volker U, Hecker M. A comprehensive two-dimensional map of cytosolic proteins of Bacillus subtilis. Electrophoresis. 2001;22:2908–2935. - PubMed
-
- Dong J, Qiu H, Garcia-Barrio M, Anderson J, Hinnebusch AG. Uncharged tRNA activates GCN2 by displacing the protein kinase moiety from a bipartite tRNA-binding domain. Mol Cell. 2000;6:269–279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
