

Targeted chiral analysis of bioactive arachidonic Acid metabolites using liquid-chromatography-mass spectrometry
- PMID: 24957514
- PMCID: PMC3901208
- DOI: 10.3390/metabo2020337
Targeted chiral analysis of bioactive arachidonic Acid metabolites using liquid-chromatography-mass spectrometry
Abstract
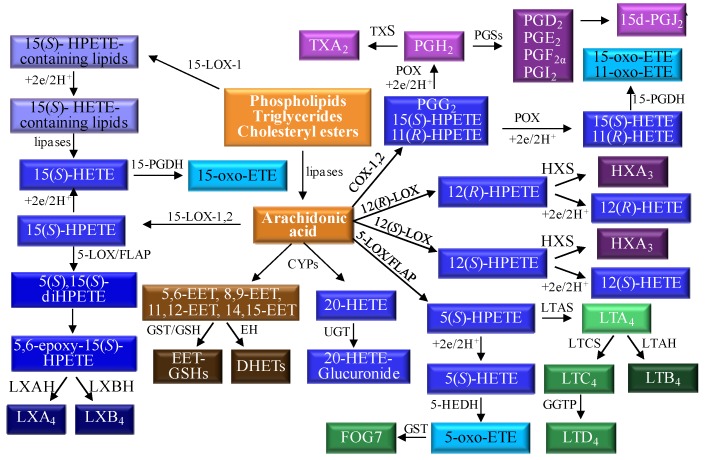
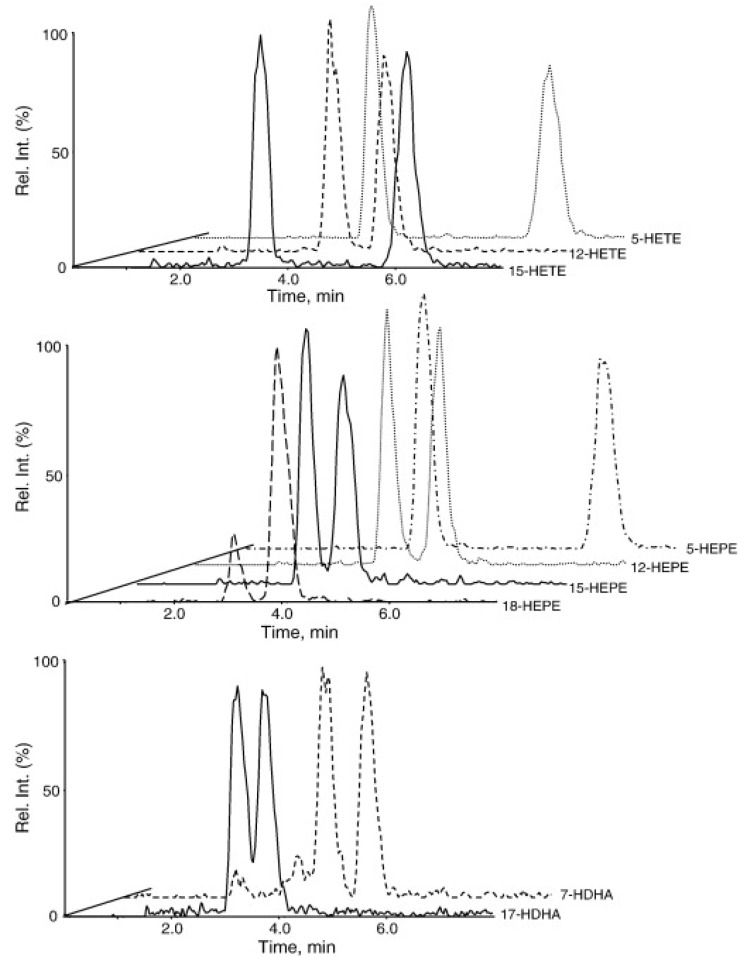
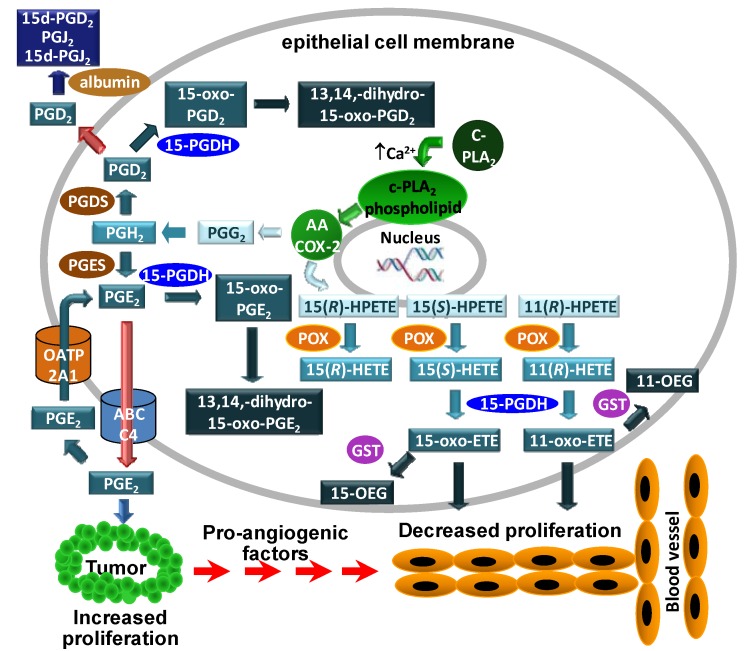
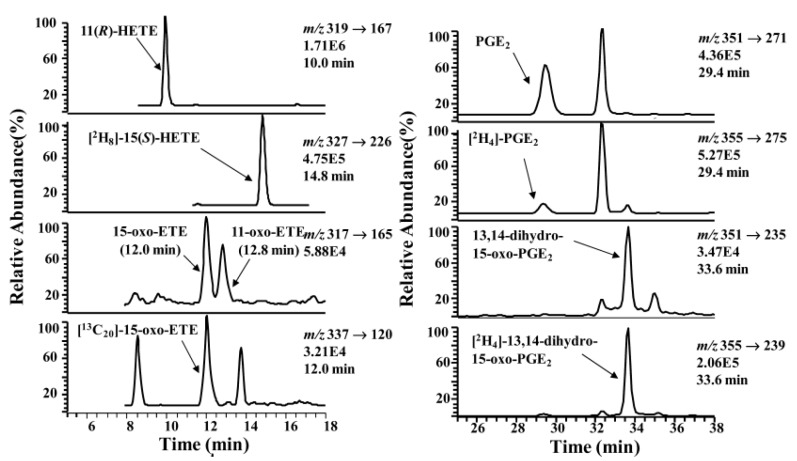
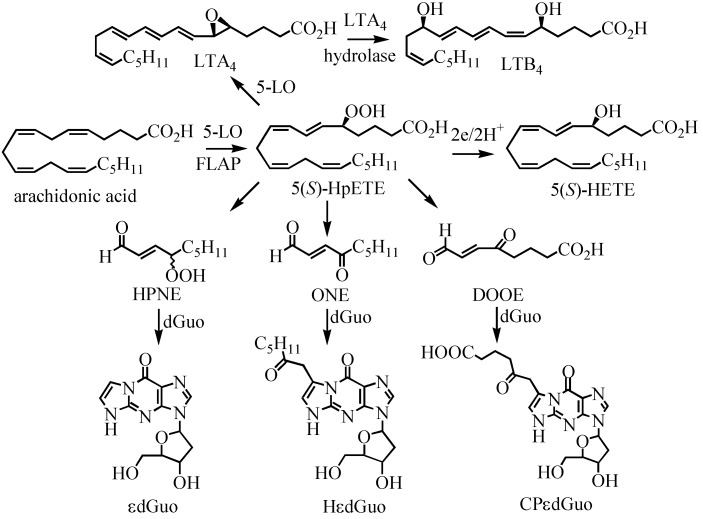
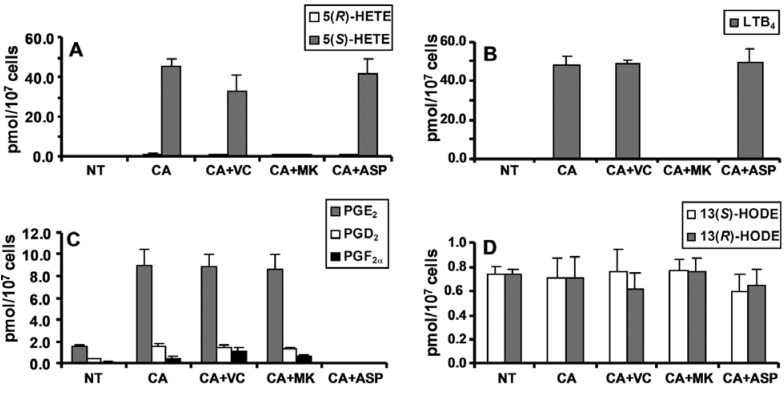
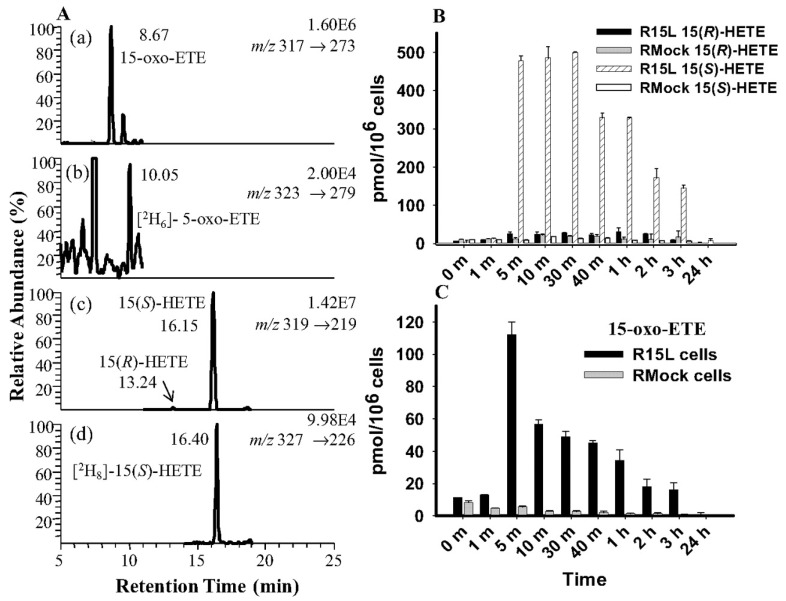
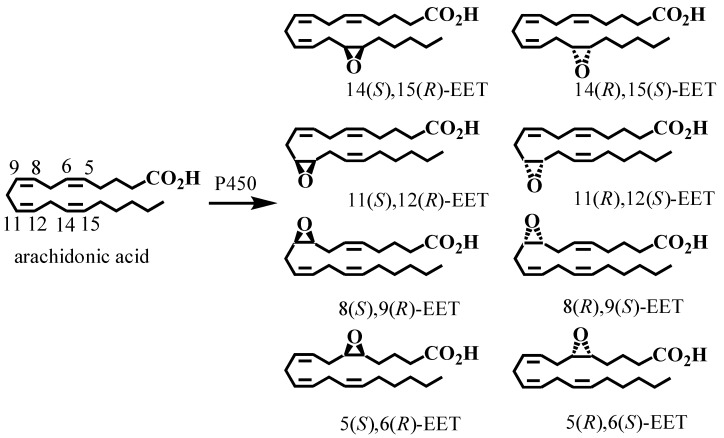
A complex structurally diverse series of eicosanoids arises from the metabolism of arachidonic acid. The metabolic profile is further complicated by the enantioselectivity of eicosanoid formation and the variety of regioisomers that arise. In order to investigate the metabolism of arachidonic acid in vitro or in vivo, targeted methods are advantageous in order to distinguish between the complex isomeric mixtures that can arise by different metabolic pathways. Over the last several years this targeted approach has become more popular, although there are still relatively few examples where chiral targeted approaches have been employed to directly analyze complex enantiomeric mixtures. To efficiently conduct targeted eicosanoid analyses, LC separations are coupled with collision induced dissociation (CID) and tandem mass spectrometry (MS/MS). Product ion profiles are often diagnostic for particular regioisomers. The highest sensitivity that can be achieved involves the use of selected reaction monitoring/mass spectrometry (SRM/MS); whereas the highest specificity is obtained with an SRM transitions between an intense parent ion, which contains the intact molecule (M) and a structurally significant product ion. This review article provides an overview of arachidonic acid metabolism and targeted chiral methods that have been utilized for the analysis of the structurally diverse eicosanoids that arise.
Figures
Similar articles
-
Targeted chiral lipidomics analysis of bioactive eicosanoid lipids in cellular systems.BMB Rep. 2009 Jul 31;42(7):401-10. doi: 10.5483/bmbrep.2009.42.7.401. BMB Rep. 2009. PMID: 19643036 Free PMC article. Review.
-
Targeted quantitative analysis of eicosanoid lipids in biological samples using liquid chromatography-tandem mass spectrometry.J Chromatogr B Analyt Technol Biomed Life Sci. 2009 Sep 15;877(26):2736-45. doi: 10.1016/j.jchromb.2009.03.011. Epub 2009 Mar 17. J Chromatogr B Analyt Technol Biomed Life Sci. 2009. PMID: 19345647 Free PMC article. Review.
-
Isobaric metabolite interferences and the requirement for close examination of raw data in addition to stringent chromatographic separations in liquid chromatography/tandem mass spectrometric analysis of drugs in biological matrix.Rapid Commun Mass Spectrom. 2008 Jul;22(13):2021-8. doi: 10.1002/rcm.3577. Rapid Commun Mass Spectrom. 2008. PMID: 18512848
-
A novel approach for quantitative peptides analysis by selected electron transfer reaction monitoring.J Chromatogr A. 2010 Oct 29;1217(44):6927-31. doi: 10.1016/j.chroma.2010.08.068. Epub 2010 Sep 17. J Chromatogr A. 2010. PMID: 20850119
-
Cellular lipid extraction for targeted stable isotope dilution liquid chromatography-mass spectrometry analysis.J Vis Exp. 2011 Nov 17;(57):3399. doi: 10.3791/3399. J Vis Exp. 2011. PMID: 22127066 Free PMC article.
Cited by
-
Identification of a Pharmacological Biomarker for the Bioassay-Based Quality Control of a Thirteen-Component TCM Formula (Lianhua Qingwen) Used in Treating Influenza A Virus (H1N1) Infection.Front Pharmacol. 2020 May 25;11:746. doi: 10.3389/fphar.2020.00746. eCollection 2020. Front Pharmacol. 2020. PMID: 32523531 Free PMC article.
-
Modern Methods of Sample Preparation for the Analysis of Oxylipins in Biological Samples.Molecules. 2019 Apr 25;24(8):1639. doi: 10.3390/molecules24081639. Molecules. 2019. PMID: 31027298 Free PMC article. Review.
-
Enantioselectivity Effects in Clinical Metabolomics and Lipidomics.Molecules. 2021 Aug 28;26(17):5231. doi: 10.3390/molecules26175231. Molecules. 2021. PMID: 34500665 Free PMC article. Review.
-
Targeted lipidomic strategies for oxygenated metabolites of polyunsaturated fatty acids.Biochim Biophys Acta. 2015 Apr;1851(4):456-68. doi: 10.1016/j.bbalip.2014.11.012. Epub 2014 Dec 5. Biochim Biophys Acta. 2015. PMID: 25486530 Free PMC article. Review.
-
Chiral lipidomics of monoepoxy and monohydroxy metabolites derived from long-chain polyunsaturated fatty acids.J Lipid Res. 2019 Jan;60(1):135-148. doi: 10.1194/jlr.M089755. Epub 2018 Nov 8. J Lipid Res. 2019. PMID: 30409844 Free PMC article.
References
-
- Bergstrom S., Samuelsson B. Isolation of prostaglandin E1 from human seminal plasma. Prostaglandins and related factors. 11. J. Biol. Chem. 1962;237:3005–3006. - PubMed
-
- Hamberg M., Samuelsson B. Oxygenation of unsaturated fatty acids by the vesicular gland of sheep. J. Biol. Chem. 1967;242:5344–5354. - PubMed
-
- DuBois R.N., Abramson S.B., Crofford L., Gupta R.A., Simon L.S., van de Putte L.B., Lipsky P.E. Cyclooxygenase in biology and disease. FASEB J. 1998;12:1063–1073. - PubMed
-
- Crofford L.J. COX-1 and COX-2 tissue expression: implications and predictions. J. Rheumatol. Suppl. 1997;49:15–19. - PubMed
-
- Pong S.S., Hong S.L., Levine L. Prostaglandin production by methylcholanthrene-transformed mouse BALB/3T3. Requirement for protein synthesis. J. Biol. Chem. 1977;252:1408–1413. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
