

Distinct HIV-1 entry phenotypes are associated with transmission, subtype specificity, and resistance to broadly neutralizing antibodies
- PMID: 24957778
- PMCID: PMC4230403
- DOI: 10.1186/1742-4690-11-48
Distinct HIV-1 entry phenotypes are associated with transmission, subtype specificity, and resistance to broadly neutralizing antibodies
Abstract
Background: The efficiency of CD4/CCR5 mediated HIV-1 entry has important implications for pathogenesis and transmission. The HIV-1 receptor affinity profiling (Affinofile) system analyzes and quantifies the infectivity of HIV-1 envelopes (Envs) across a spectrum of CD4/CCR5 expression levels and distills these data into a set of Affinofile metrics. The Affinofile system has shed light on how differential CD4/CCR5 usage efficiencies contributes to an array of Env phenotypes associated with cellular tropism, viral pathogenesis, and CCR5 inhibitor resistance. To facilitate more rapid, convenient, and robust analysis of HIV-1 entry phenotypes, we engineered a reporter Affinofile system containing a Tat- and Rev-dependent Gaussia luciferase-eGFP-Reporter (GGR) that is compatible with the use of pseudotyped or replication competent viruses with or without a virally encoded reporter gene. This GGR Affinofile system enabled a higher throughput characterization of CD4/CCR5 usage efficiencies associated with differential Env phenotypes.
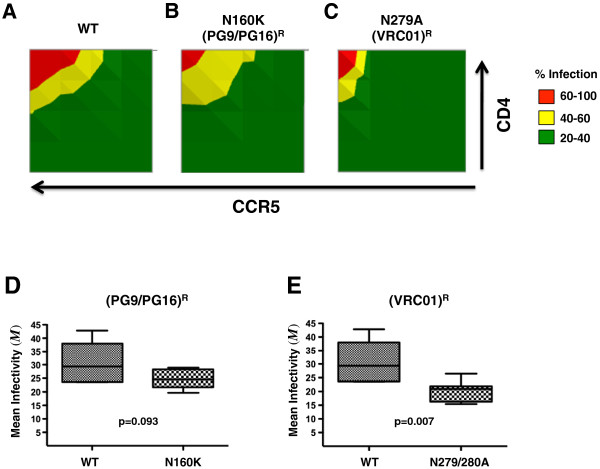
Results: We first validated our GGR Affinofile system on isogenic JR-CSF Env mutants that differ in their affinity for CD4 and/or CCR5. We established that their GGR Affinofile metrics reflected their differential entry phenotypes on primary PBMCs and CD4+ T-cell subsets. We then applied GGR Affinofile profiling to reveal distinct entry phenotypes associated with transmission, subtype specificity, and resistance to broadly neutralizing antibodies (BNAbs). First, we profiled a panel of reference subtype B transmitted/founder (T/F) and chronic Envs (n = 12) by analyzing the infectivity of each Env across 25 distinct combinations of CD4/CCR5 expression levels. Affinofile metrics revealed that at low CCR5 levels, our panel of subtype B T/F Envs was more dependent on high levels of CD4 for HIV-1 entry compared to chronic Envs. Next, we analyzed a reference panel of 28 acute/early subtype A-D Envs, and noted that subtype C Envs could be distinguished from the other subtypes based on their infectivity profiles and relevant Affinofile metrics. Lastly, mutations known to confer resistance to VRC01 or PG6/PG19 BNAbs, when engineered into subtypes A-D Envs, resulted in significantly decreased CD4/CCR5 usage efficiency.
Conclusions: GGR Affinofile profiling reveals pathophysiological phenotypes associated with varying HIV-1 entry efficiencies, and highlight the fitness costs associated with resistance to some broadly neutralizing antibodies.
Figures






References
-
- Wilen CB, Tilton JC, Doms RW. Molecular mechanisms of HIV entry. Adv Exp Med Biol. 2012;726:223–242. - PubMed
-
- Spira S, Wainberg M, Loemba H, Turner D, Brenner B. Impact of clade diversity on HIV-1 virulence, antiretroviral drug sensitivity and drug resistance. J Antimicrob Chemoth. 2003;51:229–240. - PubMed
-
- Gorry PR, Ancuta P. Coreceptors and HIV-1 Pathogenesis. Curr HIV/AIDS Rep. 2010;8:45–53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
