

Small molecule inhibitors of HCV replication from pomegranate
- PMID: 24958333
- PMCID: PMC4067622
- DOI: 10.1038/srep05411
Small molecule inhibitors of HCV replication from pomegranate
Abstract
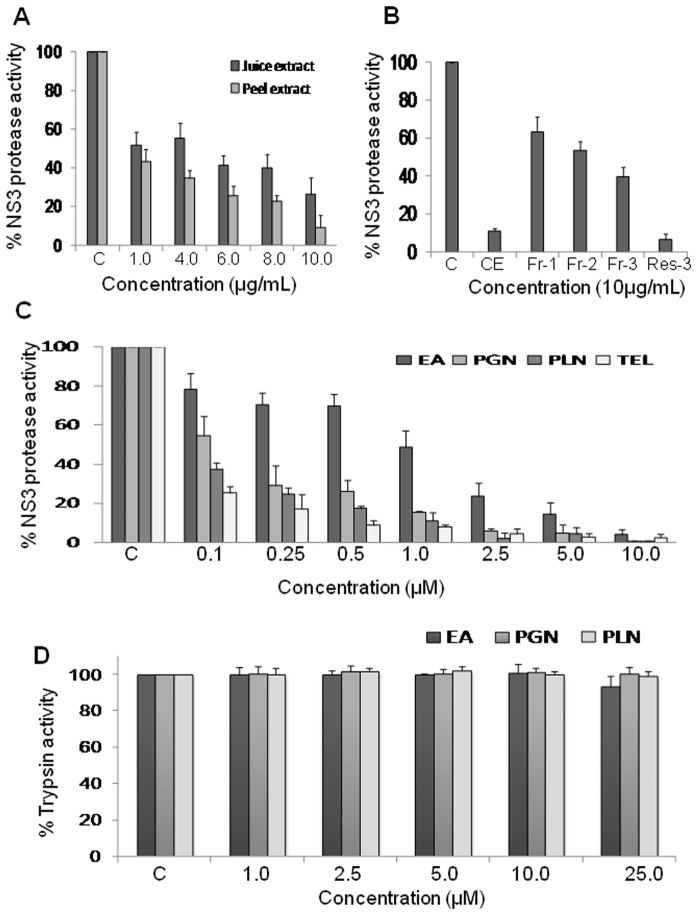
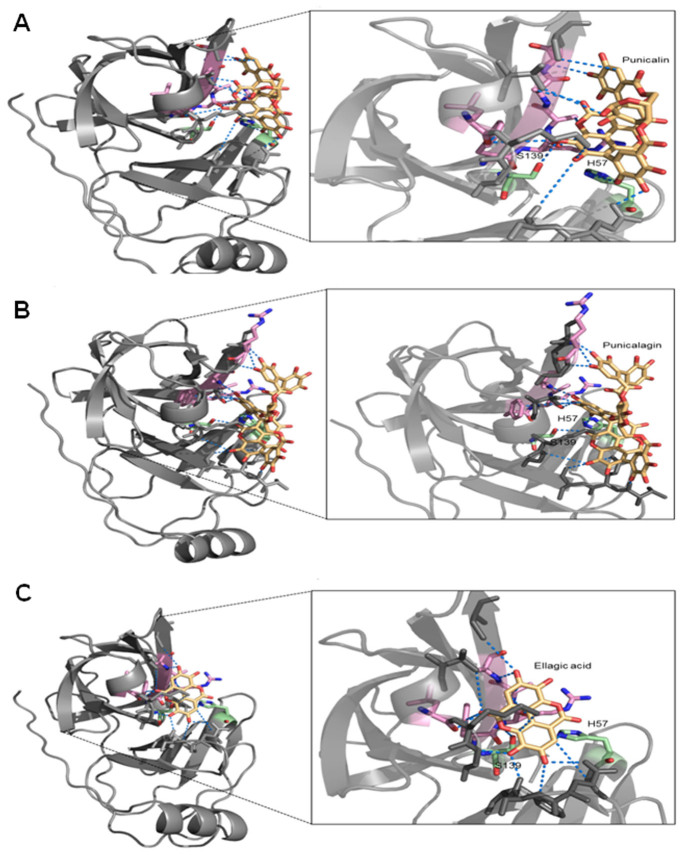
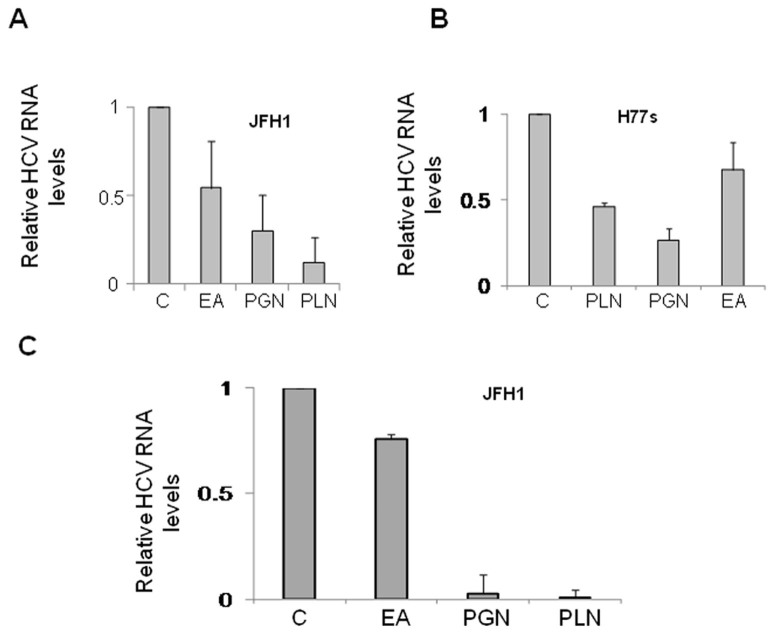
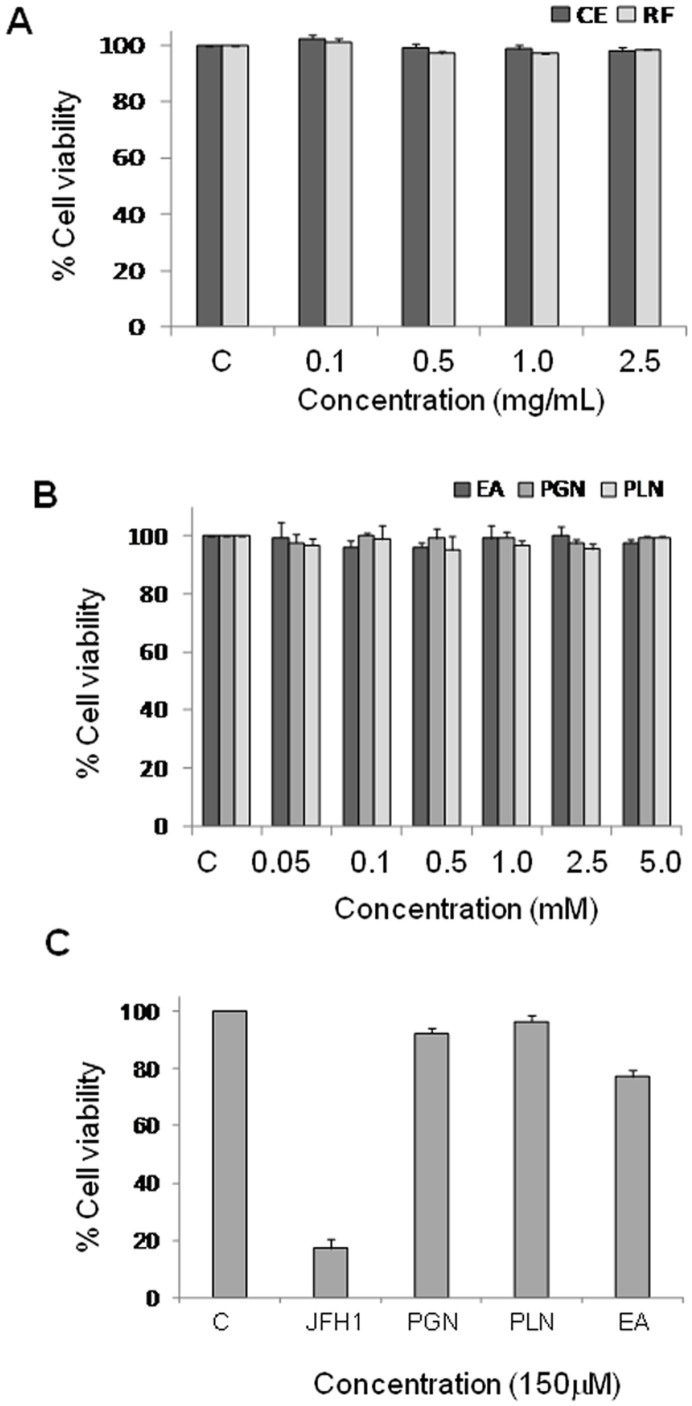
Hepatitis C virus (HCV) is the causative agent of end-stage liver disease. Recent advances in the last decade in anti HCV treatment strategies have dramatically increased the viral clearance rate. However, several limitations are still associated, which warrant a great need of novel, safe and selective drugs against HCV infection. Towards this objective, we explored highly potent and selective small molecule inhibitors, the ellagitannins, from the crude extract of Pomegranate (Punica granatum) fruit peel. The pure compounds, punicalagin, punicalin, and ellagic acid isolated from the extract specifically blocked the HCV NS3/4A protease activity in vitro. Structural analysis using computational approach also showed that ligand molecules interact with the catalytic and substrate binding residues of NS3/4A protease, leading to inhibition of the enzyme activity. Further, punicalagin and punicalin significantly reduced the HCV replication in cell culture system. More importantly, these compounds are well tolerated ex vivo and'no observed adverse effect level' (NOAEL) was established upto an acute dose of 5000 mg/kg in BALB/c mice. Additionally, pharmacokinetics study showed that the compounds are bioavailable. Taken together, our study provides a proof-of-concept approach for the potential use of antiviral and non-toxic principle ellagitannins from pomegranate in prevention and control of HCV induced complications.
Figures
References
-
- Boyer N. & Marcellin P. Pathogenesis, diagnosis and management of hepatitis C. J. Hepatol. 32, 98–112 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
