

Intestinal epithelium-specific MyD88 signaling impacts host susceptibility to infectious colitis by promoting protective goblet cell and antimicrobial responses
- PMID: 24958710
- PMCID: PMC4187802
- DOI: 10.1128/IAI.02045-14
Intestinal epithelium-specific MyD88 signaling impacts host susceptibility to infectious colitis by promoting protective goblet cell and antimicrobial responses
Abstract
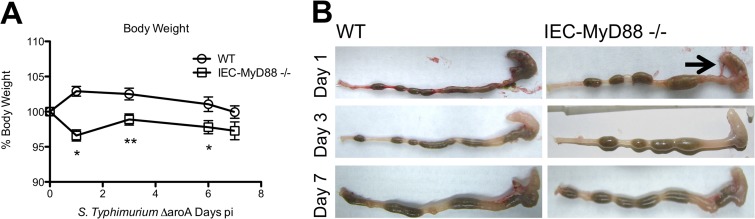
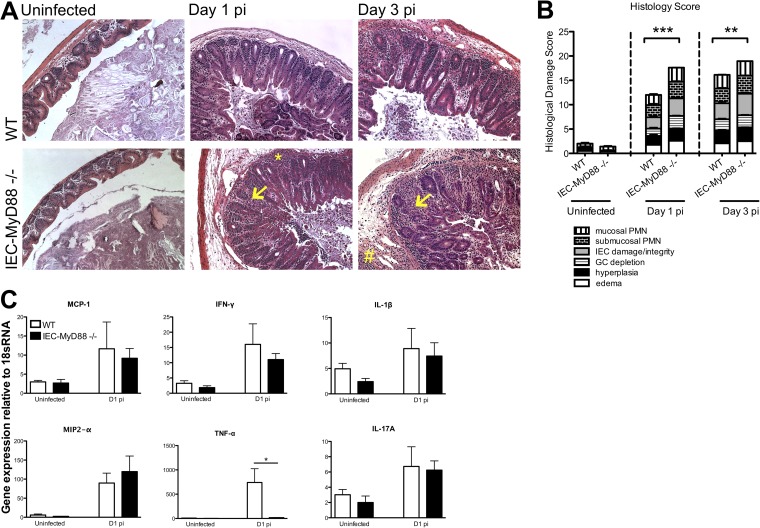
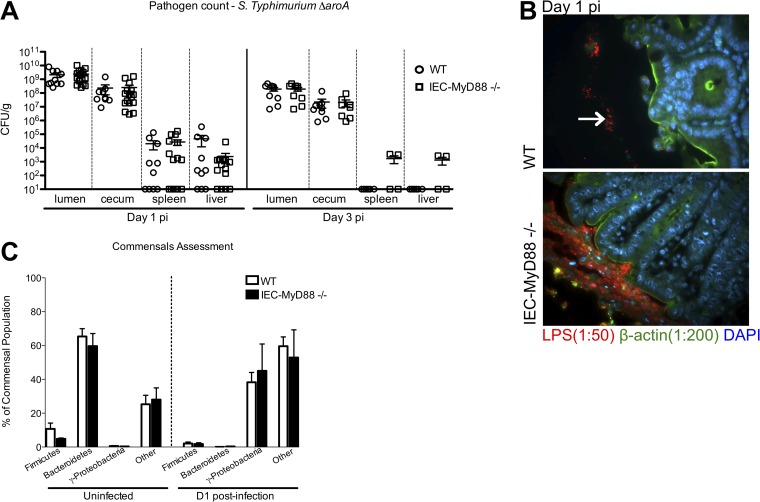
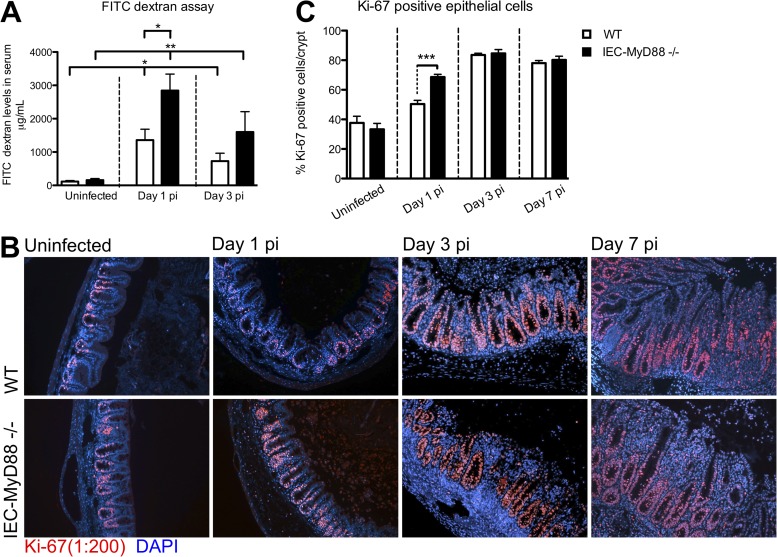
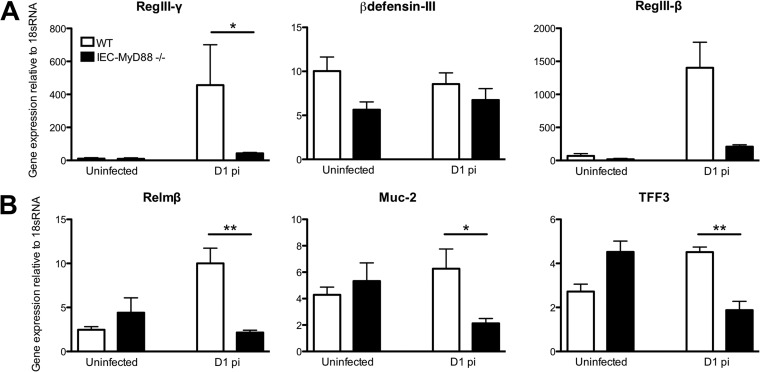
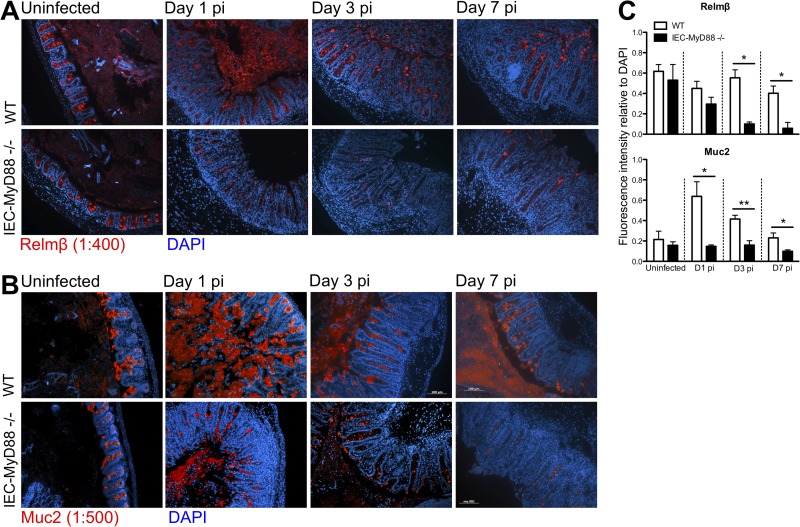
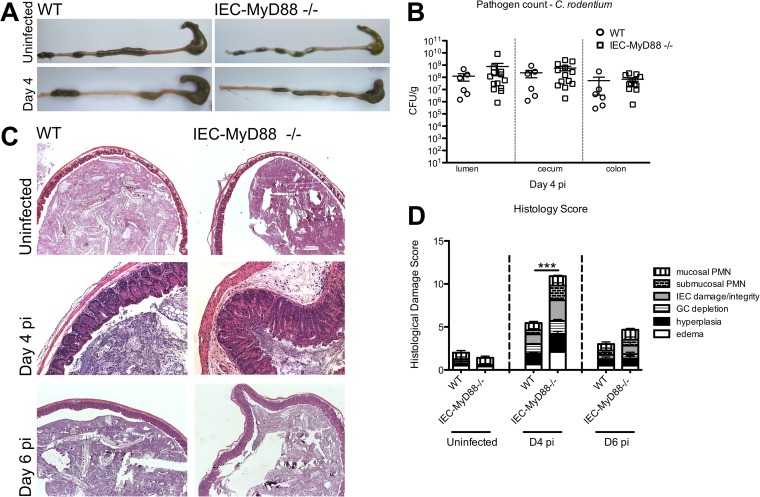
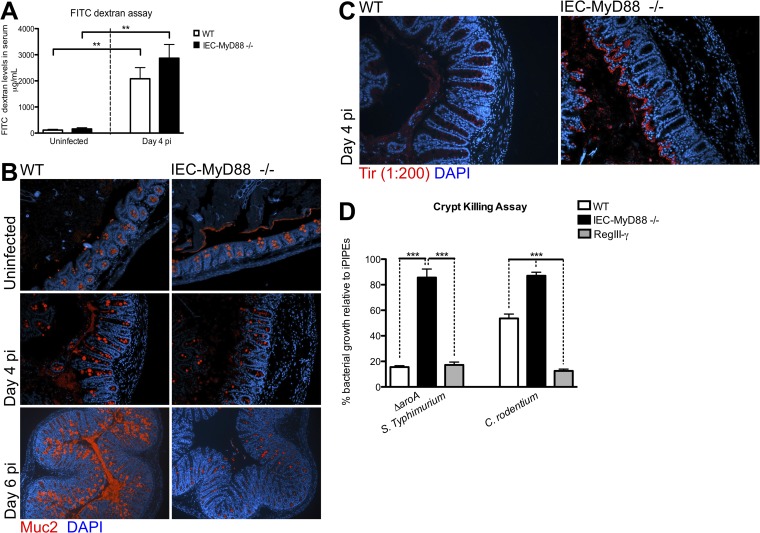
Intestinal epithelial cells (IECs), including secretory goblet cells, form essential physiochemical barriers that separate luminal bacteria from underlying immune cells in the intestinal mucosa. IECs are common targets for enteric bacterial pathogens, with hosts responding to these microbes through innate toll-like receptors that predominantly signal through the MyD88 adaptor protein. In fact, MyD88 signaling confers protection against several enteric bacterial pathogens, including Salmonella enterica serovar Typhimurium and Citrobacter rodentium. Since IECs are considered innately hyporesponsive, it is unclear whether MyD88 signaling within IECs contributes to this protection. We infected mice lacking MyD88 solely in their IECs (IEC-Myd88(-/-)) with S. Typhimurium. Compared to wild-type (WT) mice, infected IEC-Myd88(-/-) mice suffered accelerated tissue damage, exaggerated barrier disruption, and impaired goblet cell responses (Muc2 and RELMβ). Immunostaining revealed S. Typhimurium penetrated the IECs of IEC-Myd88(-/-) mice, unlike in WT mice, where they were sequestered to the lumen. When isolated crypts were assayed for their antimicrobial actions, crypts from IEC-Myd88(-/-) mice were severely impaired in their antimicrobial activity against S. Typhimurium. We also examined whether MyD88 signaling in IECs impacted host defense against C. rodentium, with IEC-Myd88(-/-) mice again suffering exaggerated tissue damage, impaired goblet cell responses, and reduced antimicrobial activity against C. rodentium. These results demonstrate that MyD88 signaling within IECs plays an important protective role at early stages of infection, influencing host susceptibility to infection by controlling the ability of the pathogen to reach and survive at the intestinal mucosal surface.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Gibson DL, Ma C, Bergstrom KSB, Huang JT, Man C, Vallance BA. 2008. MyD88 signalling plays a critical role in host defence by controlling pathogen burden and promoting epithelial cell homeostasis during Citrobacter rodentium-induced colitis. Cell. Microbiol. 10:618–631. 10.1111/j.1462-5822.2007.01071.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
