

Piezo1, a mechanically activated ion channel, is required for vascular development in mice
- PMID: 24958852
- PMCID: PMC4104881
- DOI: 10.1073/pnas.1409233111
Piezo1, a mechanically activated ion channel, is required for vascular development in mice
Abstract
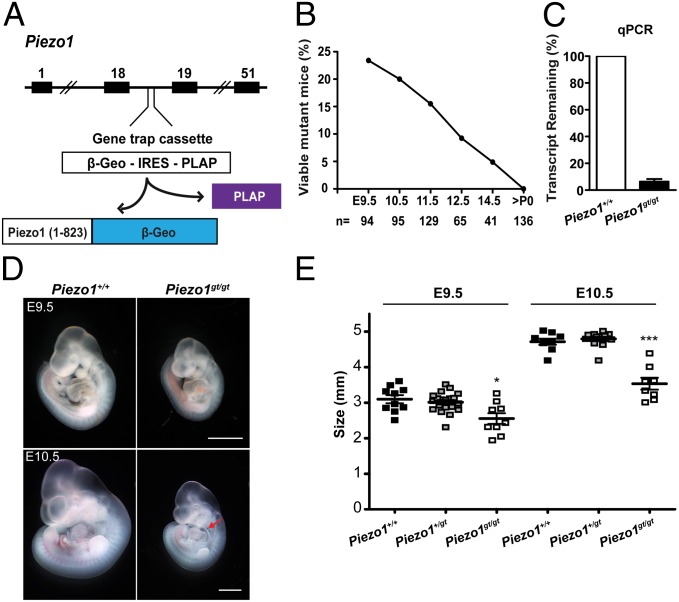
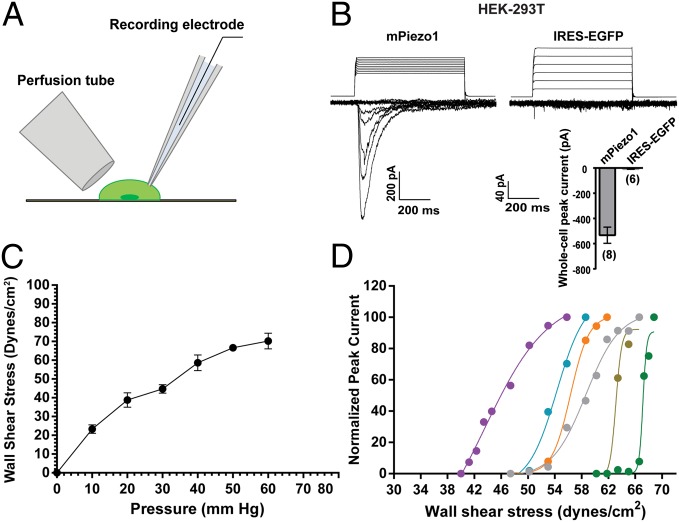
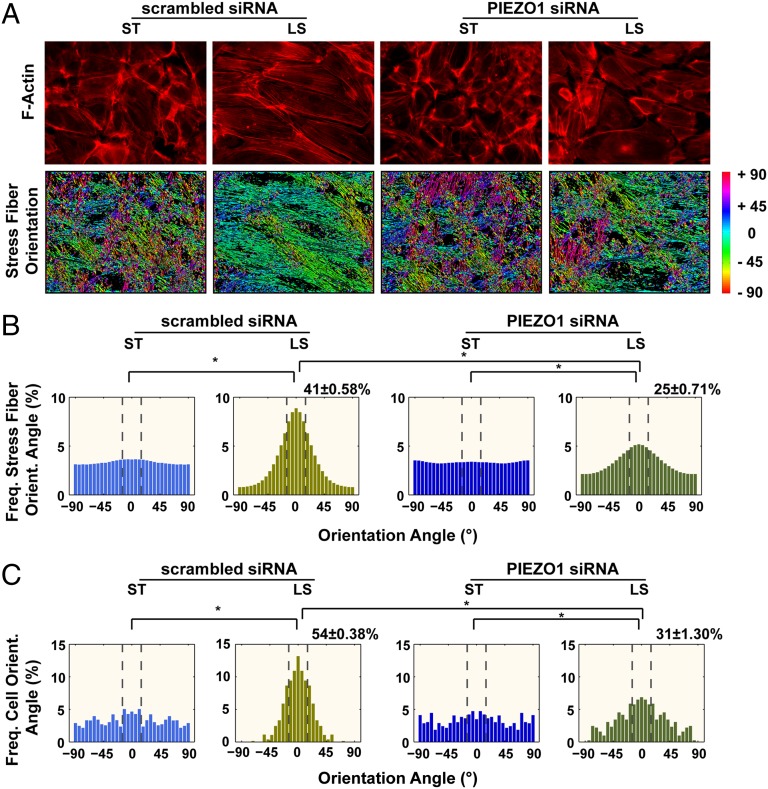
Mechanosensation is perhaps the last sensory modality not understood at the molecular level. Ion channels that sense mechanical force are postulated to play critical roles in a variety of biological processes including sensing touch/pain (somatosensation), sound (hearing), and shear stress (cardiovascular physiology); however, the identity of these ion channels has remained elusive. We previously identified Piezo1 and Piezo2 as mechanically activated cation channels that are expressed in many mechanosensitive cell types. Here, we show that Piezo1 is expressed in endothelial cells of developing blood vessels in mice. Piezo1-deficient embryos die at midgestation with defects in vascular remodeling, a process critically influenced by blood flow. We demonstrate that Piezo1 is activated by shear stress, the major type of mechanical force experienced by endothelial cells in response to blood flow. Furthermore, loss of Piezo1 in endothelial cells leads to deficits in stress fiber and cellular orientation in response to shear stress, linking Piezo1 mechanotransduction to regulation of cell morphology. These findings highlight an essential role of mammalian Piezo1 in vascular development during embryonic development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Orr AW, Helmke BP, Blackman BR, Schwartz MA. Mechanisms of mechanotransduction. Dev Cell. 2006;10(1):11–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
