

Haptoglobin is required to prevent oxidative stress and muscle atrophy
- PMID: 24959824
- PMCID: PMC4069100
- DOI: 10.1371/journal.pone.0100745
Haptoglobin is required to prevent oxidative stress and muscle atrophy
Abstract
Background: Oxidative stress (OS) plays a major role on tissue function. Several catabolic or stress conditions exacerbate OS, inducing organ deterioration. Haptoglobin (Hp) is a circulating acute phase protein, produced by liver and adipose tissue, and has an important anti-oxidant function. Hp is induced in pro-oxidative conditions such as systemic inflammation or obesity. The role of systemic factors that modulate oxidative stress inside muscle cells is still poorly investigated.
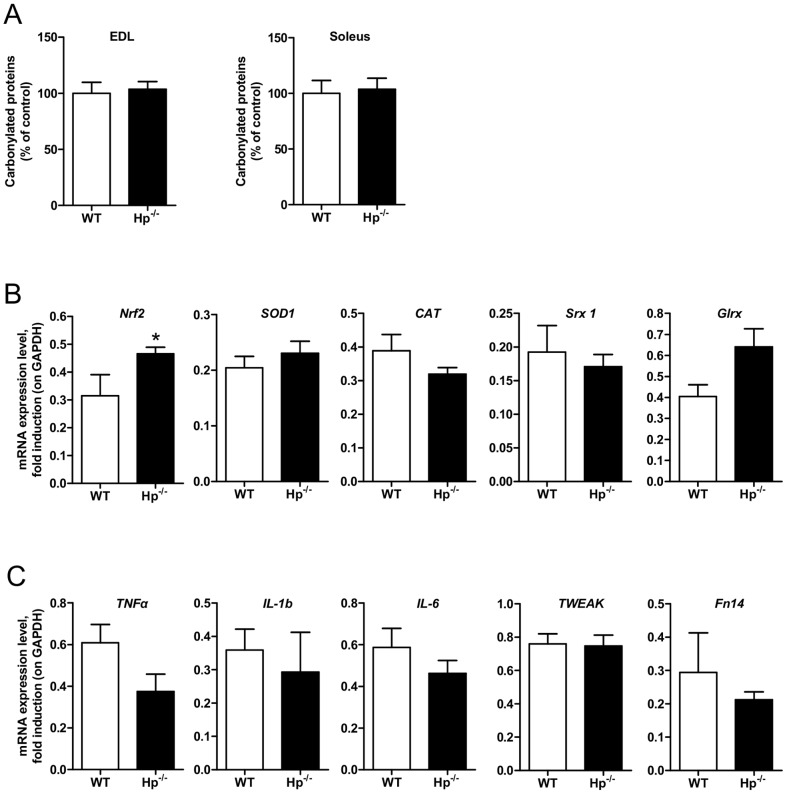
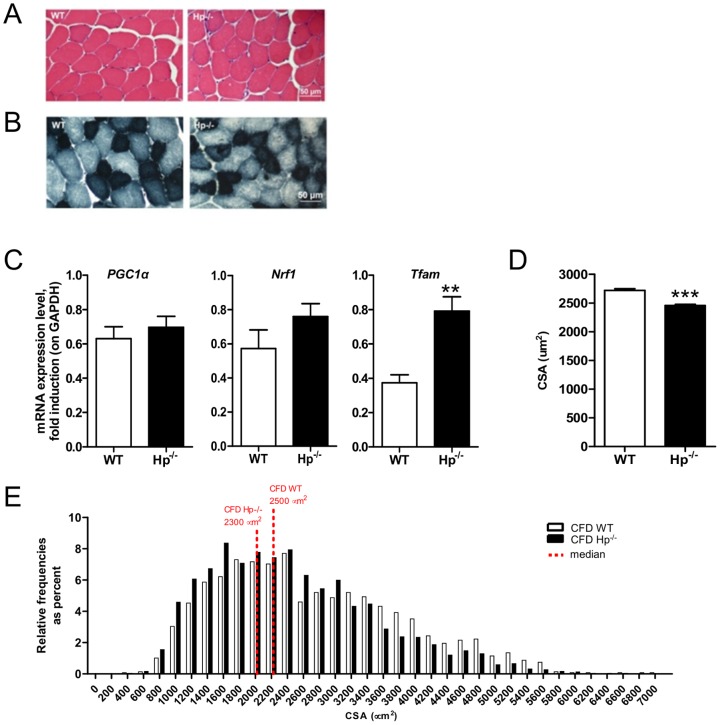
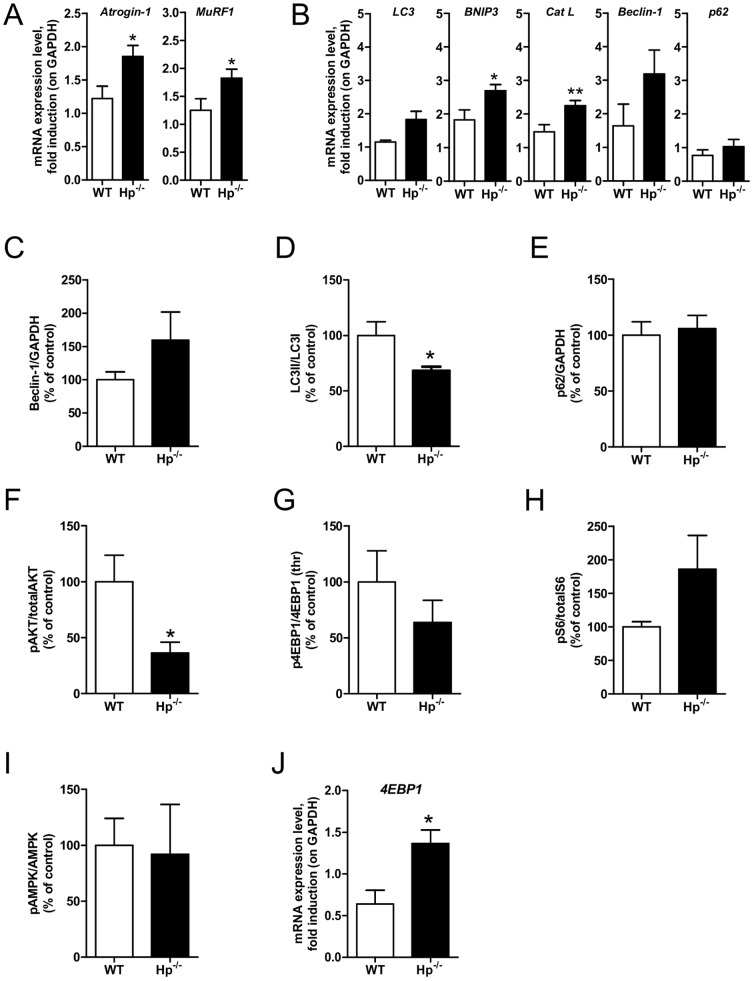
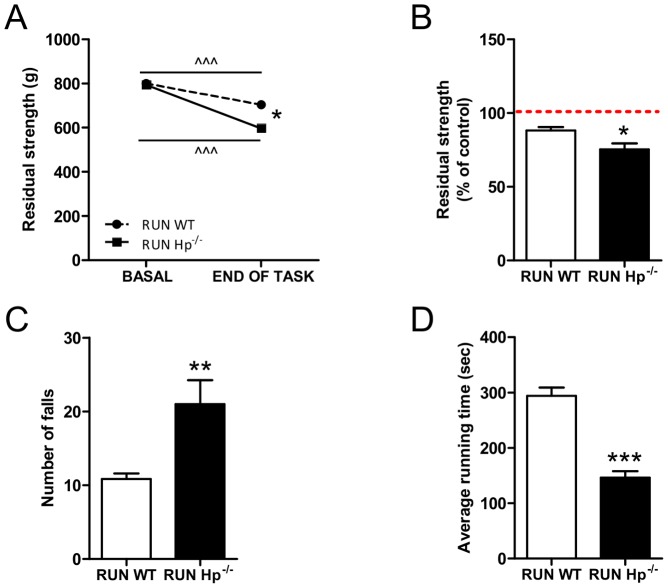
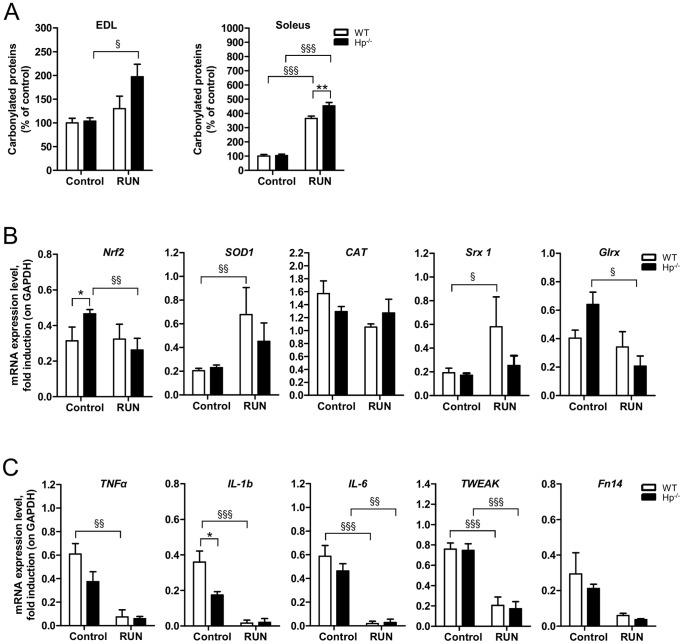
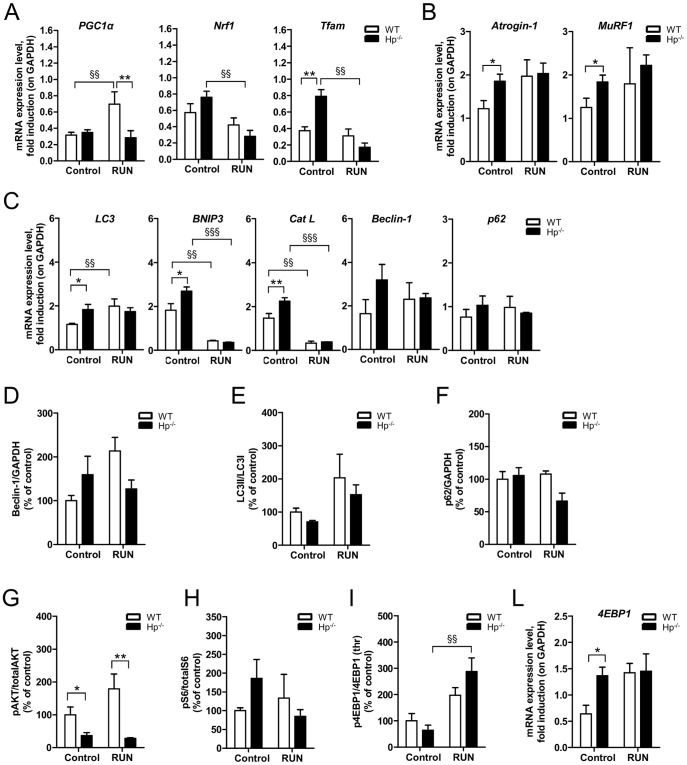
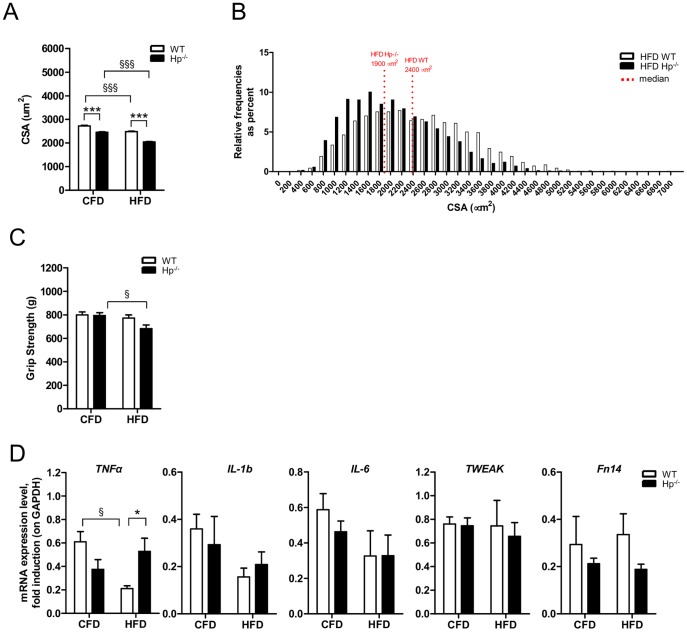
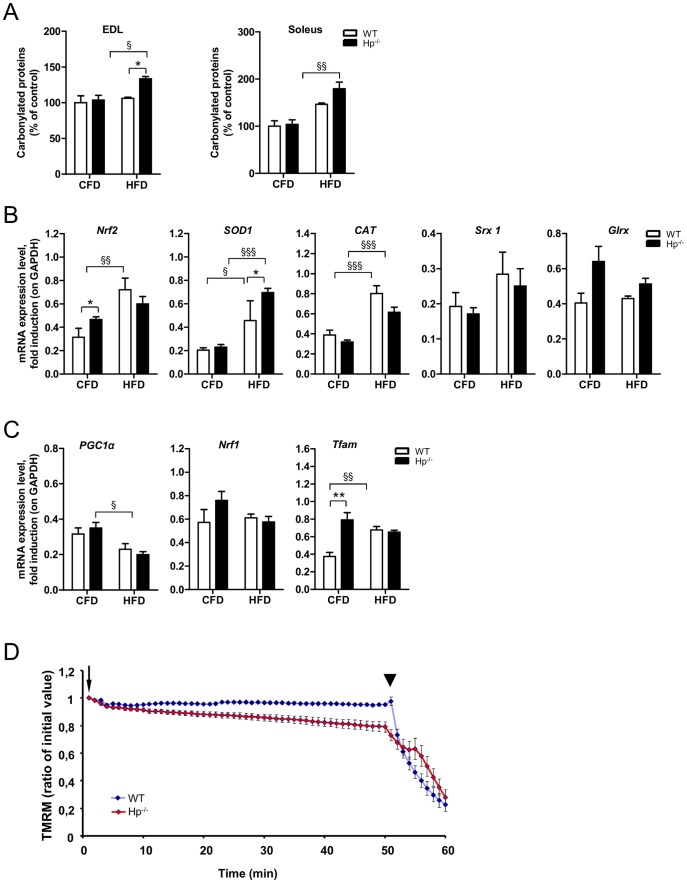
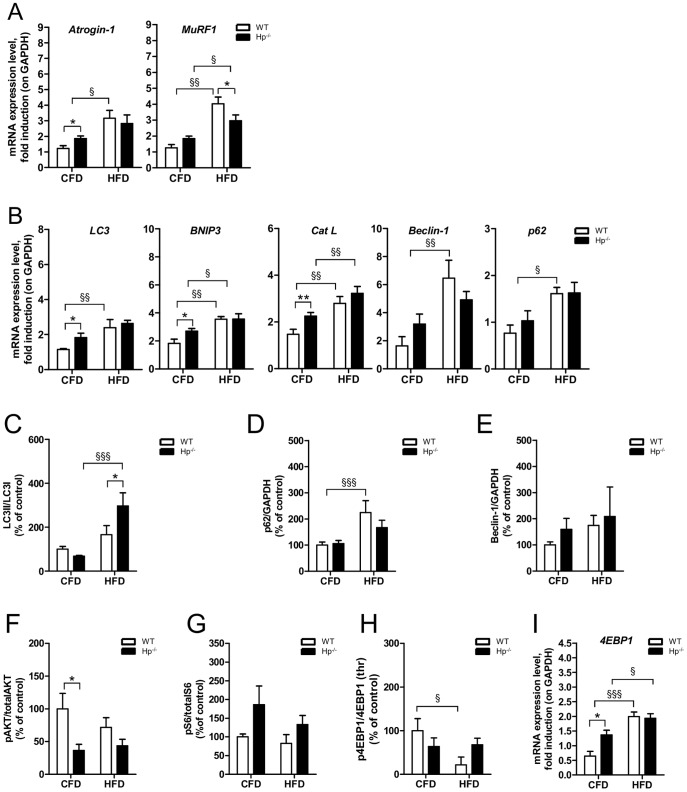
Results: We used Hp knockout mice (Hp-/-) to determine the role of this protein and therefore, of systemic OS in maintenance of muscle mass and function. Absence of Hp caused muscle atrophy and weakness due to activation of an atrophy program. When animals were stressed by acute exercise or by high fat diet (HFD), OS, muscle atrophy and force drop were exacerbated in Hp-/-. Depending from the stress condition, autophagy-lysosome and ubiquitin-proteasome systems were differently induced.
Conclusions: Hp is required to prevent OS and the activation of pathways leading to muscle atrophy and weakness in normal condition and upon metabolic challenges.
Conflict of interest statement
Figures
References
-
- Dobryszycka W (1997) Biological functions of haptoglobin—new pieces to an old puzzle. Eur J Clin Chem Clin Biochem 35: 647–654. - PubMed
-
- Chiellini C, Bertacca A, Novelli SE, Gorgun CZ, Ciccarone A, et al. (2002) Obesity modulates the expression of haptoglobin in the white adipose tissue via TNFalpha. J Cell Physiol 190: 251–258. - PubMed
-
- Kristiansen M, Graversen JH, Jacobsen C, Sonne O, Hoffman HJ, et al. (2001) Identification of the haemoglobin scavenger receptor. Nature 409: 198–201. - PubMed
-
- Lim SK, Kim H, Lim SK, bin Ali A, Lim YK, et al. (1998) Increased susceptibility in Hp knockout mice during acute hemolysis. Blood 92: 1870–1877. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
