

Wnt Signaling in Neurogenesis during Aging and Physical Activity
- PMID: 24961268
- PMCID: PMC4061808
- DOI: 10.3390/brainsci2040745
Wnt Signaling in Neurogenesis during Aging and Physical Activity
Abstract
Over the past decade, much progress has been made regarding our understanding of neurogenesis in both young and old animals and where it occurs throughout the lifespan, although the growth of new neurons declines with increasing age. In addition, physical activity can reverse this age-dependent decline in neurogenesis. Highly correlated with this decline is the degree of inter and intracellular Wnt signaling, the molecular mechanisms of which have only recently started to be elucidated. So far, most of what we know about intracellular signaling during/following exercise centers around the CREB/CRE initiated transcriptional events. Relatively little is known, however, about how aging and physical activity affect the Wnt signaling pathway. Herein, we briefly review the salient features of neurogenesis in young and then in old adult animals. Then, we discuss Wnt signaling and review the very few in vitro and in vivo studies that have examined the Wnt signaling pathways in aging and physical activity.
Figures
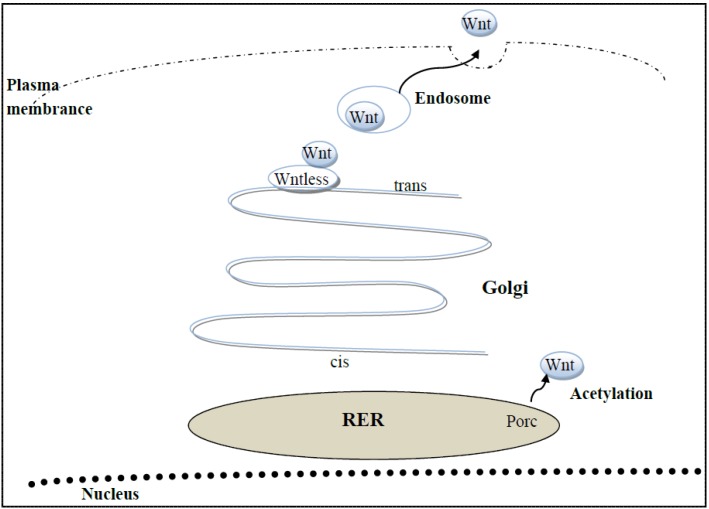
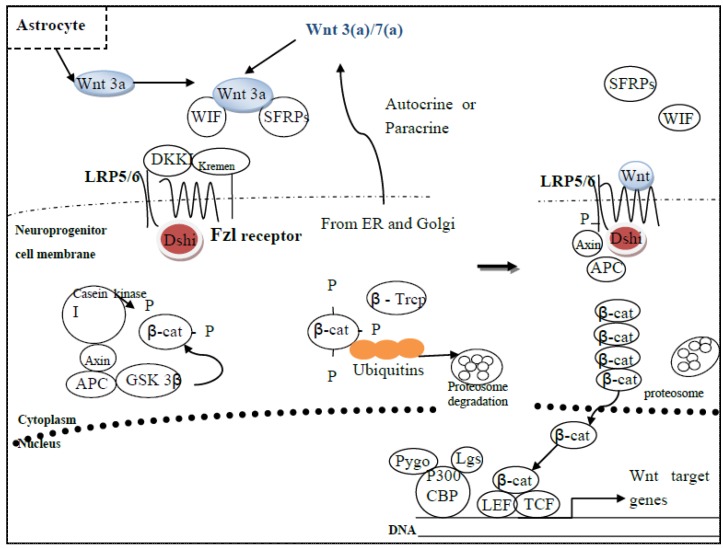
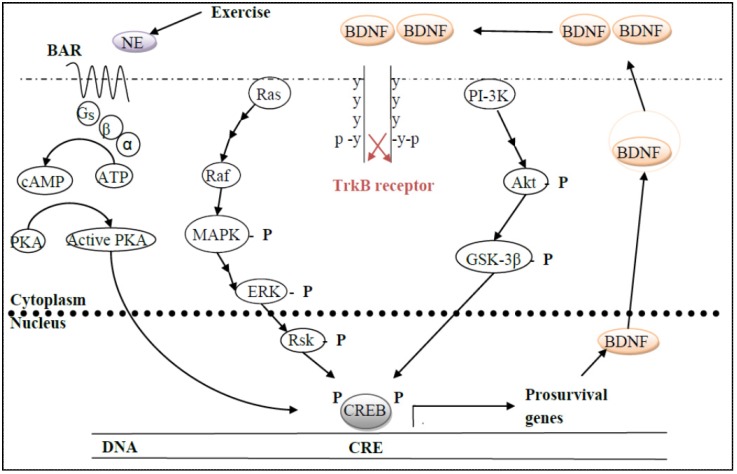
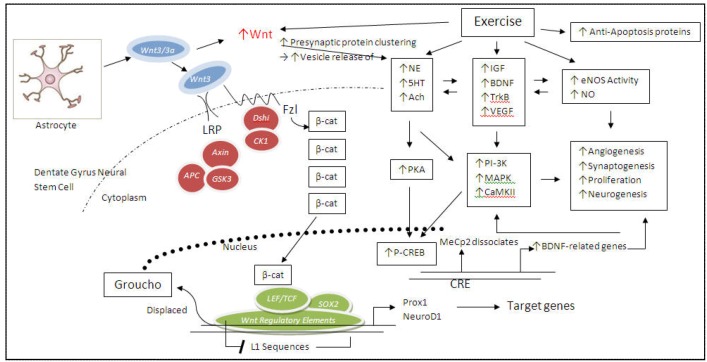
References
LinkOut - more resources
Full Text Sources
Miscellaneous
