

The roles and regulation of multicellular rosette structures during morphogenesis
- PMID: 24961796
- PMCID: PMC4067956
- DOI: 10.1242/dev.101444
The roles and regulation of multicellular rosette structures during morphogenesis
Abstract
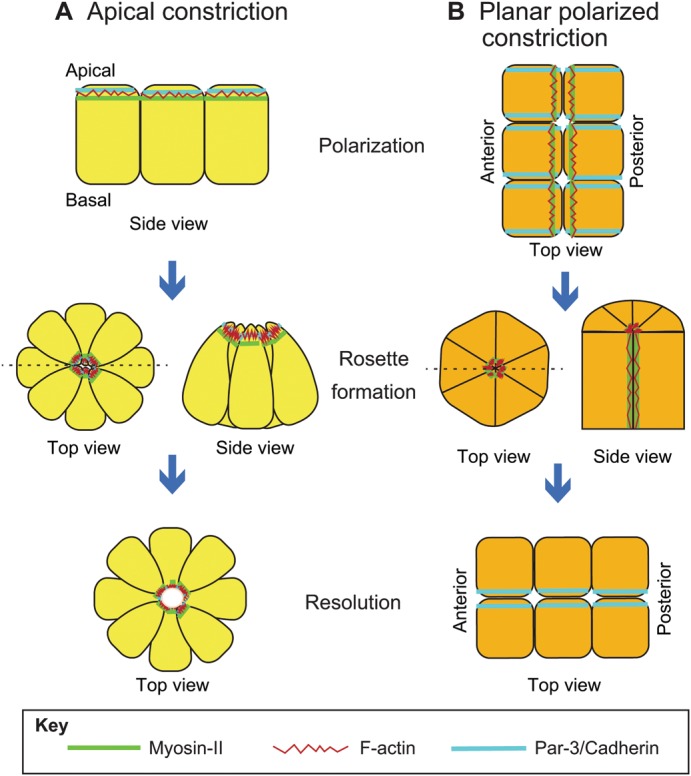
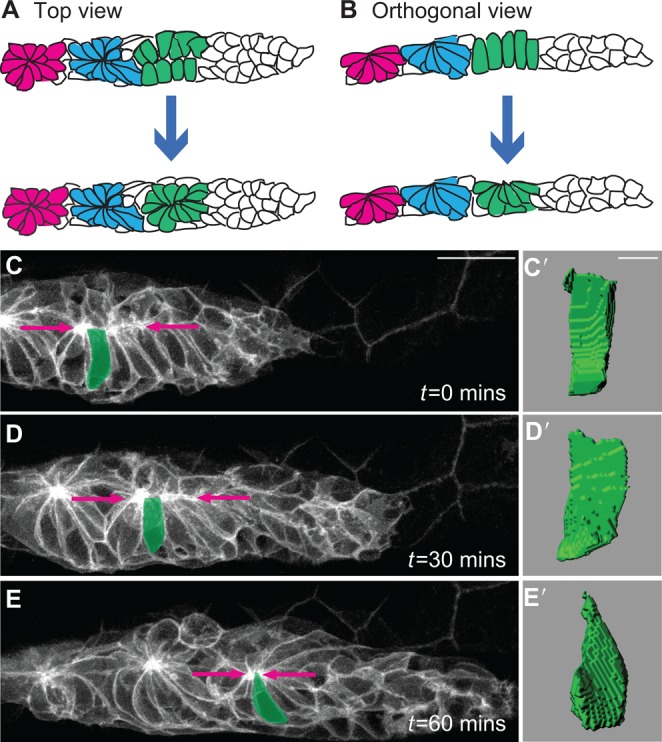
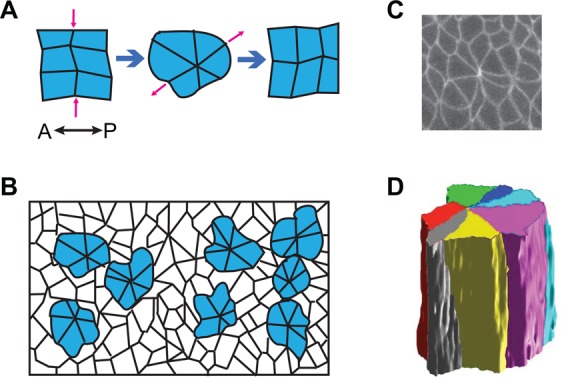
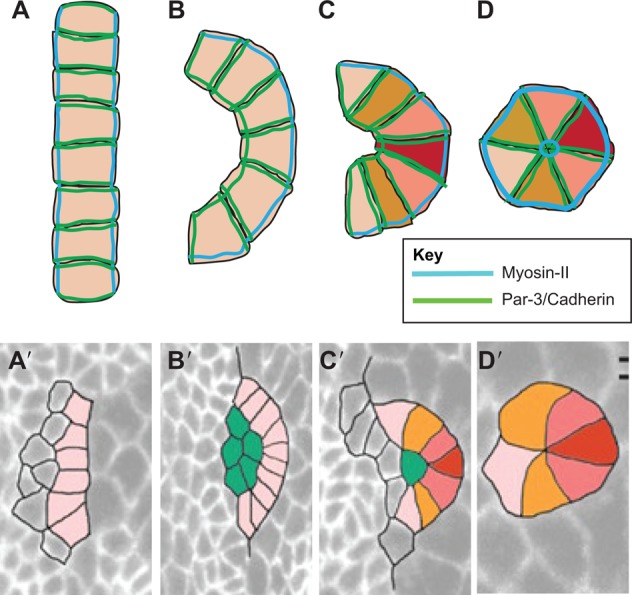
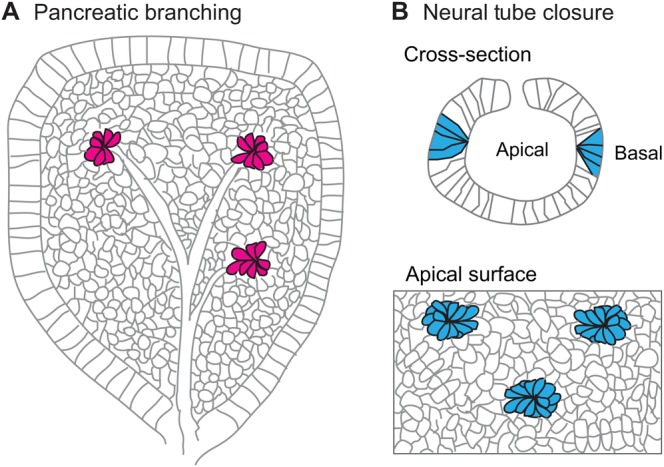
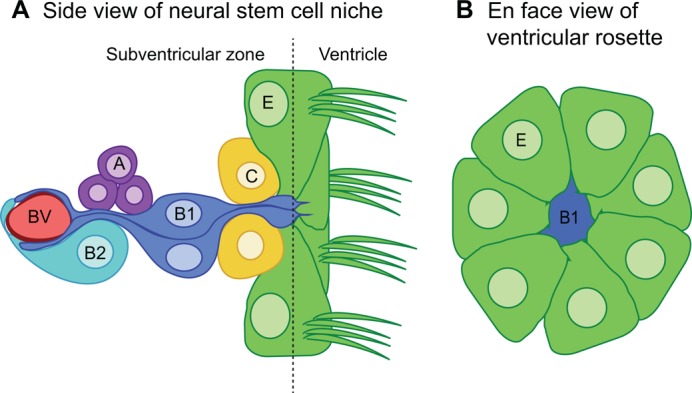
Multicellular rosettes have recently been appreciated as important cellular intermediates that are observed during the formation of diverse organ systems. These rosettes are polarized, transient epithelial structures that sometimes recapitulate the form of the adult organ. Rosette formation has been studied in various developmental contexts, such as in the zebrafish lateral line primordium, the vertebrate pancreas, the Drosophila epithelium and retina, as well as in the adult neural stem cell niche. These studies have revealed that the cytoskeletal rearrangements responsible for rosette formation appear to be conserved. By contrast, the extracellular cues that trigger these rearrangements in vivo are less well understood and are more diverse. Here, we review recent studies of the genetic regulation and cellular transitions involved in rosette formation. We discuss and compare specific models for rosette formation and highlight outstanding questions in the field.
Keywords: Drosophila epithelium; Morphogenesis; Myosin II; Rosette; Zebrafish lateral line.
© 2014. Published by The Company of Biologists Ltd.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
