

eIF4F-like complexes formed by cap-binding homolog TbEIF4E5 with TbEIF4G1 or TbEIF4G2 are implicated in post-transcriptional regulation in Trypanosoma brucei
- PMID: 24962368
- PMCID: PMC4105752
- DOI: 10.1261/rna.045534.114
eIF4F-like complexes formed by cap-binding homolog TbEIF4E5 with TbEIF4G1 or TbEIF4G2 are implicated in post-transcriptional regulation in Trypanosoma brucei
Abstract
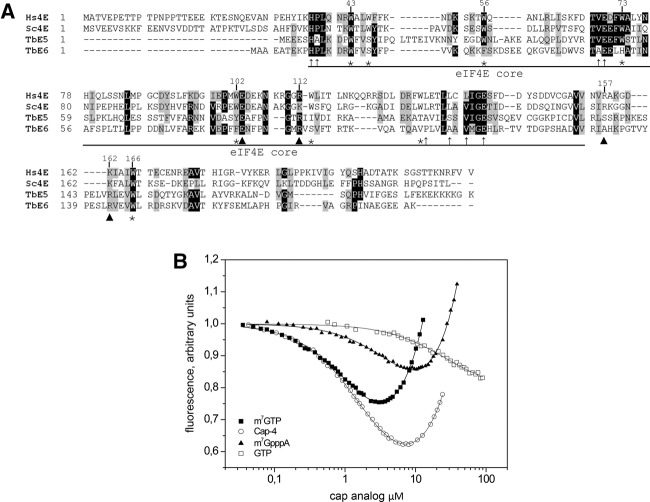
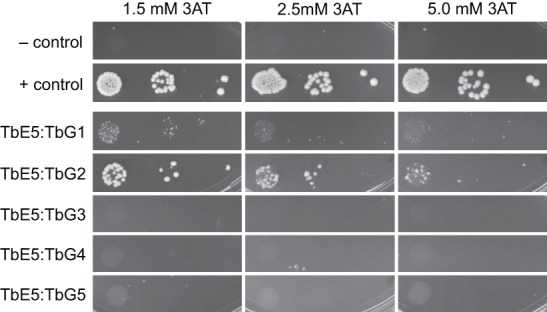
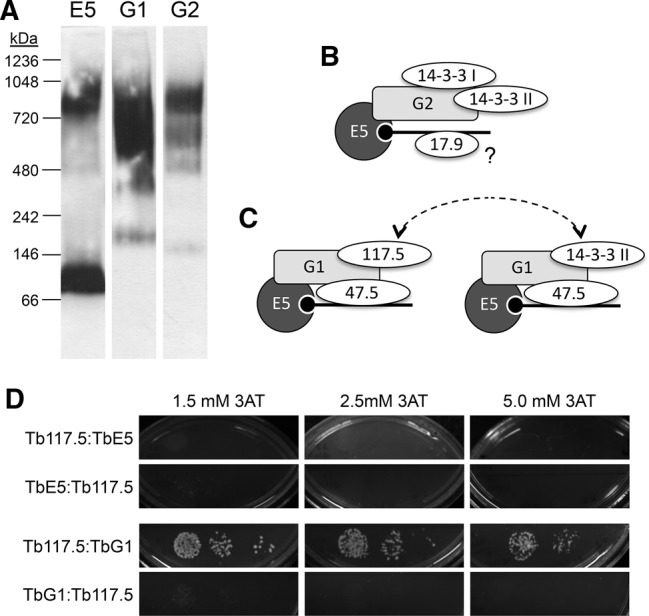
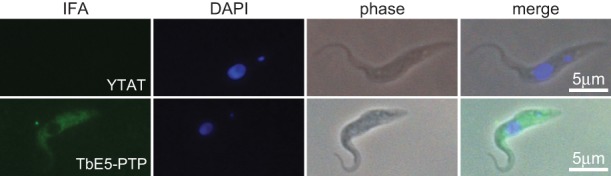
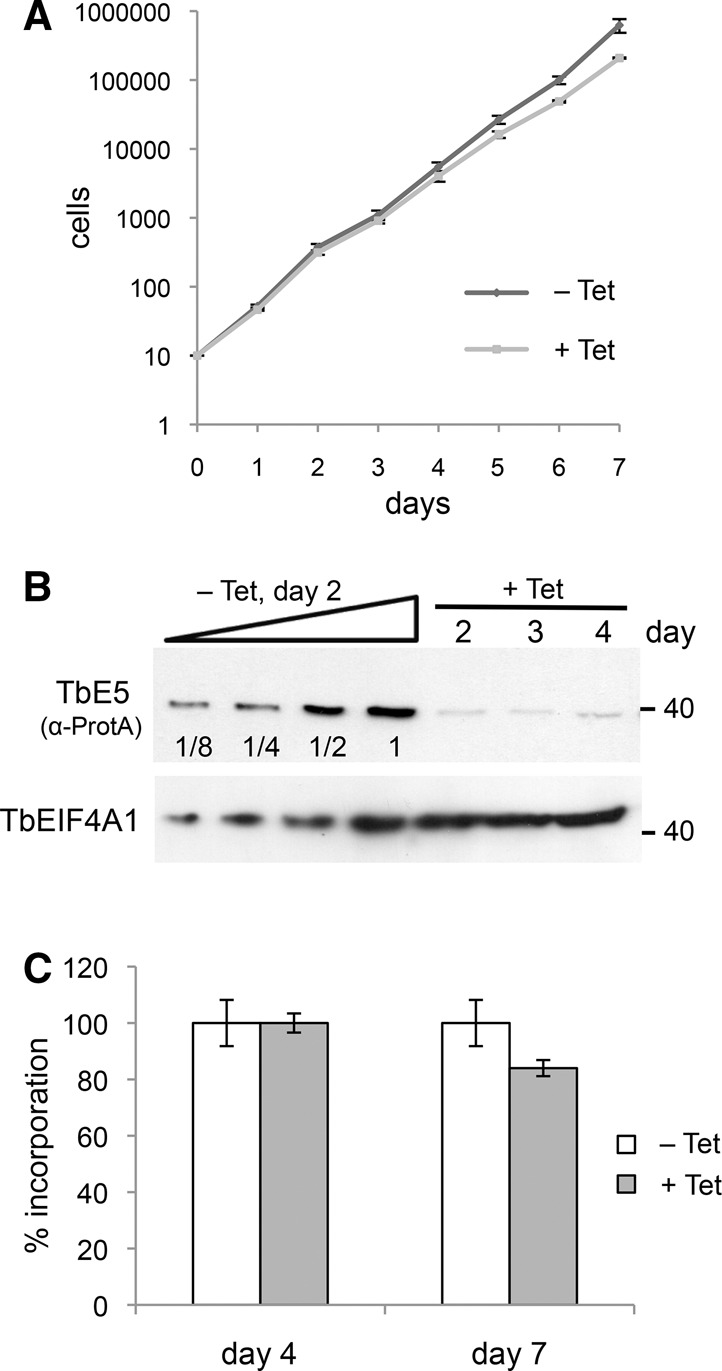
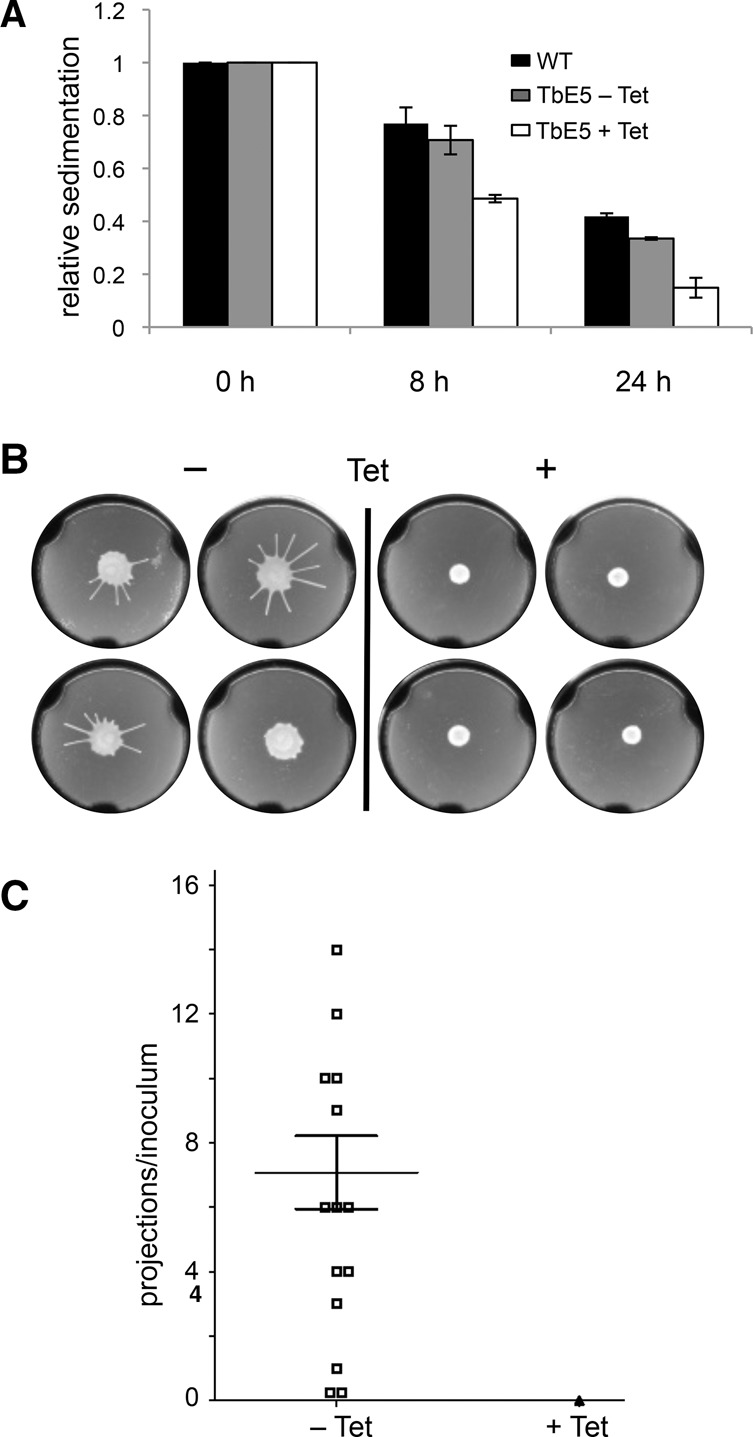
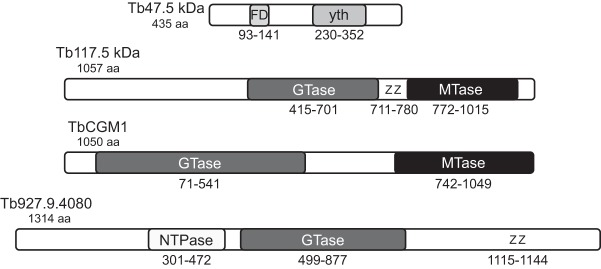
Members of the eIF4E mRNA cap-binding family are involved in translation and the modulation of transcript availability in other systems as part of a three-component complex including eIF4G and eIF4A. The kinetoplastids possess four described eIF4E and five eIF4G homologs. We have identified two new eIF4E family proteins in Trypanosoma brucei, and define distinct complexes associated with the fifth member, TbEIF4E5. The cytosolic TbEIF4E5 protein binds cap 0 in vitro. TbEIF4E5 was found in association with two of the five TbEIF4Gs. TbIF4EG1 bound TbEIF4E5, a 47.5-kDa protein with two RNA-binding domains, and either the regulatory protein 14-3-3 II or a 117.5-kDa protein with guanylyltransferase and methyltransferase domains in a potentially dynamic interaction. The TbEIF4G2/TbEIF4E5 complex was associated with a 17.9-kDa hypothetical protein and both 14-3-3 variants I and II. Knockdown of TbEIF4E5 resulted in the loss of productive cell movement, as evidenced by the inability of the cells to remain in suspension in liquid culture and the loss of social motility on semisolid plating medium, as well as a minor reduction of translation. Cells appeared lethargic, as opposed to compromised in flagellar function per se. The minimal use of transcriptional control in kinetoplastids requires these organisms to implement downstream mechanisms to regulate gene expression, and the TbEIF4E5/TbEIF4G1/117.5-kDa complex in particular may be a key player in that process. We suggest that a pathway involved in cell motility is affected, directly or indirectly, by one of the TbEIF4E5 complexes.
Keywords: 14-3-3 protein; kinetoplastid; mRNA cap; social motility; translation initiation factor.
© 2014 Freire et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
Trypanosoma brucei translation initiation factor homolog EIF4E6 forms a tripartite cytosolic complex with EIF4G5 and a capping enzyme homolog.Eukaryot Cell. 2014 Jul;13(7):896-908. doi: 10.1128/EC.00071-14. Epub 2014 May 16. Eukaryot Cell. 2014. PMID: 24839125 Free PMC article.
-
Two related trypanosomatid eIF4G homologues have functional differences compatible with distinct roles during translation initiation.RNA Biol. 2015;12(3):305-19. doi: 10.1080/15476286.2015.1017233. RNA Biol. 2015. PMID: 25826663 Free PMC article.
-
Trypanosoma brucei EIF4E2 cap-binding protein binds a homolog of the histone-mRNA stem-loop-binding protein.Curr Genet. 2018 Aug;64(4):821-839. doi: 10.1007/s00294-017-0795-3. Epub 2017 Dec 29. Curr Genet. 2018. PMID: 29288414
-
Manipulation of the host translation initiation complex eIF4F by DNA viruses.Biochem Soc Trans. 2010 Dec;38(6):1511-6. doi: 10.1042/BST0381511. Biochem Soc Trans. 2010. PMID: 21118117 Review.
-
The Role of Cytoplasmic mRNA Cap-Binding Protein Complexes in Trypanosoma brucei and Other Trypanosomatids.Pathogens. 2017 Oct 27;6(4):55. doi: 10.3390/pathogens6040055. Pathogens. 2017. PMID: 29077018 Free PMC article. Review.
Cited by
-
Characterization of an Atypical eIF4E Ortholog in Leishmania, LeishIF4E-6.Int J Mol Sci. 2021 Nov 24;22(23):12720. doi: 10.3390/ijms222312720. Int J Mol Sci. 2021. PMID: 34884522 Free PMC article.
-
Crystal structure of the Trypanosoma cruzi EIF4E5 translation factor homologue in complex with mRNA cap-4.Nucleic Acids Res. 2019 Jun 20;47(11):5973-5987. doi: 10.1093/nar/gkz339. Nucleic Acids Res. 2019. PMID: 31066441 Free PMC article.
-
The eIF3 complex of Trypanosoma brucei: composition conservation does not imply the conservation of structural assembly and subunits function.RNA. 2017 Mar;23(3):333-345. doi: 10.1261/rna.058651.116. Epub 2016 Dec 8. RNA. 2017. PMID: 27932584 Free PMC article.
-
The eIF3 complex of Leishmania-subunit composition and mode of recruitment to different cap-binding complexes.Nucleic Acids Res. 2015 Jul 27;43(13):6222-35. doi: 10.1093/nar/gkv564. Epub 2015 Jun 19. Nucleic Acids Res. 2015. PMID: 26092695 Free PMC article.
-
Structural analysis of the Trypanosoma brucei EIF4E6/EIF4G5 complex reveals details of the interaction between unusual eIF4F subunits.Sci Rep. 2024 Jan 25;14(1):2178. doi: 10.1038/s41598-024-52364-1. Sci Rep. 2024. PMID: 38272944 Free PMC article.
References
-
- Aitken CE, Lorsch JR 2012. A mechanistic overview of translation initiation in eukaryotes. Nat Struct Mol Biol 19: 568–576 - PubMed
-
- Alsford S, duBois K, Horn D, Field MC 2012. Epigenetic mechanisms, nuclear architecture and the control of gene expression in trypanosomes. Expert Rev Mol Med 14: e13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous