

Substrate specificity of cytoplasmic N-glycosyltransferase
- PMID: 24962585
- PMCID: PMC4148877
- DOI: 10.1074/jbc.M114.579326
Substrate specificity of cytoplasmic N-glycosyltransferase
Abstract
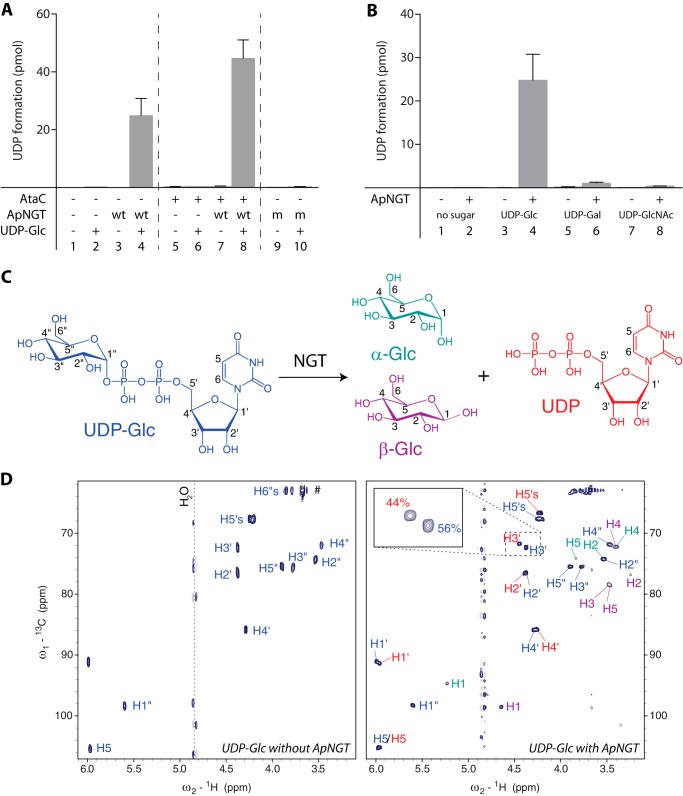
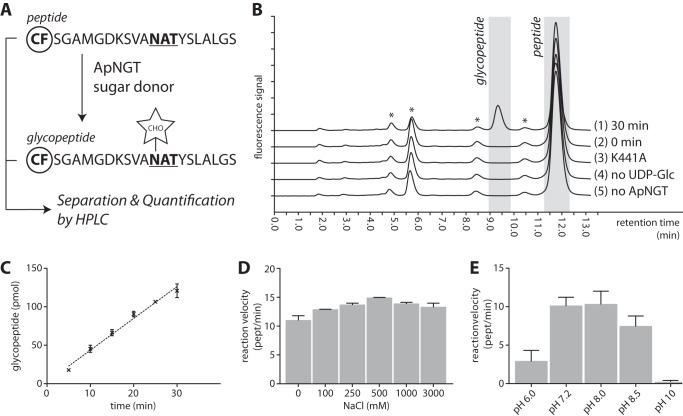
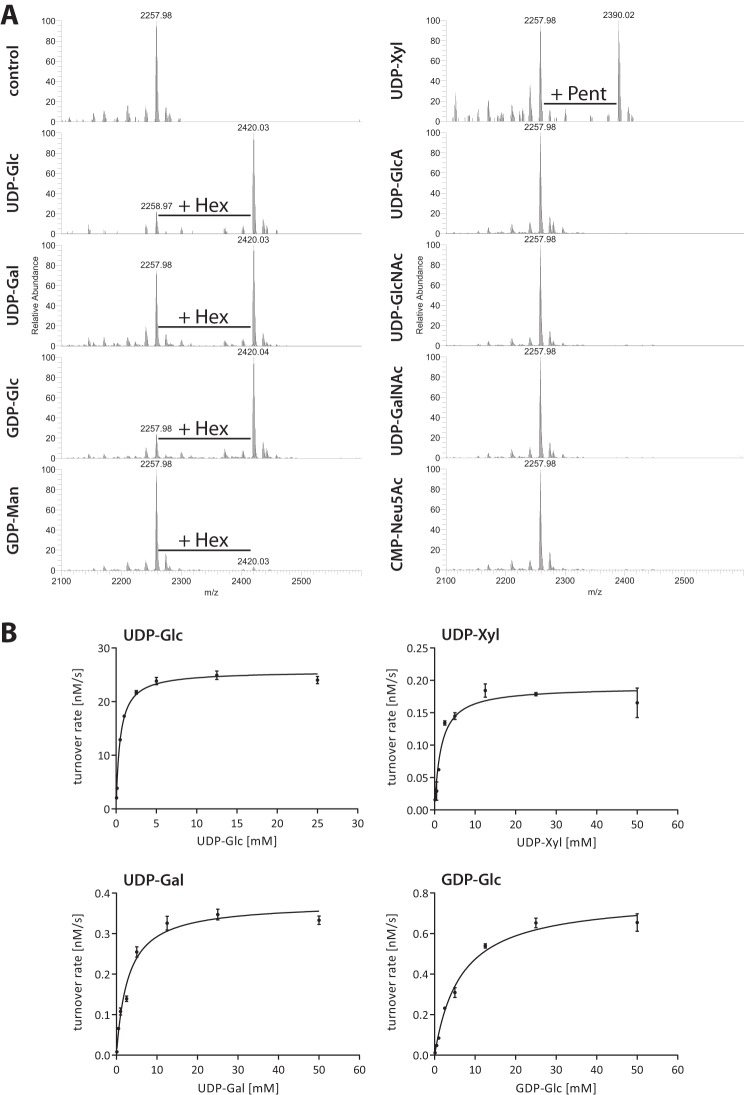
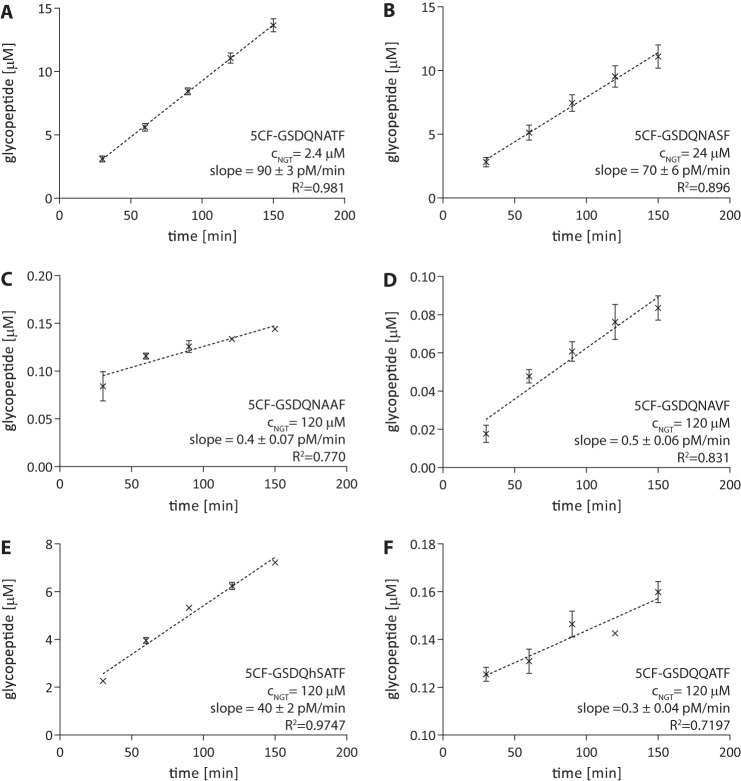
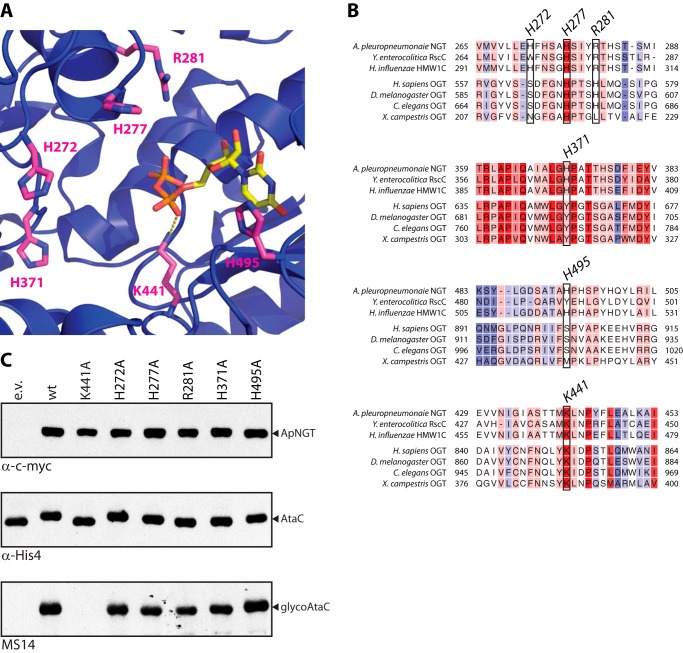
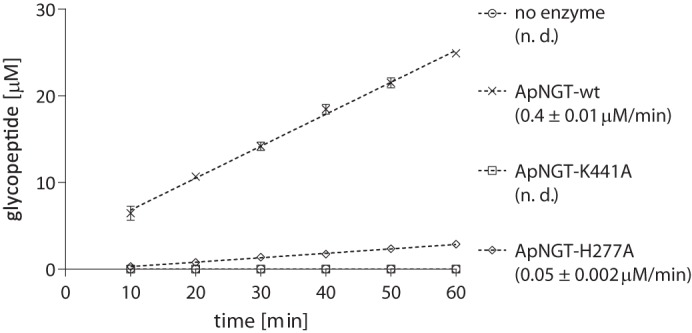
N-Linked protein glycosylation is a very common post-translational modification that can be found in all kingdoms of life. The classical, highly conserved pathway entails the assembly of a lipid-linked oligosaccharide and its transfer to an asparagine residue in the sequon NX(S/T) of a secreted protein by the integral membrane protein oligosaccharyltransferase. A few species in the class of γ-proteobacteria encode a cytoplasmic N-glycosylation system mediated by a soluble N-glycosyltransferase (NGT). This enzyme uses nucleotide-activated sugars to modify asparagine residues with single monosaccharides. As these enzymes are not related to oligosaccharyltransferase, NGTs constitute a novel class of N-glycosylation catalyzing enzymes. To characterize the NGT-catalyzed reaction, we developed a sensitive and quantitative in vitro assay based on HPLC separation and quantification of fluorescently labeled substrate peptides. With this assay we were able to directly quantify glycopeptide formation by Actinobacillus pleuropneumoniae NGT and determine its substrate specificities: NGT turns over a number of different sugar donor substrates and allows for activation by both UDP and GDP. Quantitative analysis of peptide substrate turnover demonstrated a strikingly similar specificity as the classical, oligosaccharyltransferase-catalyzed N-glycosylation, with NX(S/T) sequons being the optimal NGT substrates.
Keywords: Actinobacillus Pleuropneumoniae; Bacteria; Glycosylation; Glycosyltransferase; Post-translational Modification (PTM); Substrate Specificity.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Apweiler R., Hermjakob H., Sharon N. (1999) On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim. Biophys. Acta 1473, 4–8 - PubMed
-
- Aebi M., Bernasconi R., Clerc S., Molinari M. (2010) N-Glycan structures: recognition and processing in the ER. Trends Biochem. Sci. 35, 74–82 - PubMed
-
- Helenius A., Aebi M. (2004) Roles of N-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem. 73, 1019–1049 - PubMed
-
- Sharon N., Lis H. (1995) Lectins–proteins with a sweet tooth: functions in cell recognition. Essays Biochem. 30, 59–75 - PubMed
-
- Kaneko Y., Nimmerjahn F., Ravetch J. V. (2006) Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 313, 670–673 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
