

Characterization of Cardiac Anoctamin1 Ca²⁺-Activated Chloride Channels and Functional Role in Ischemia-Induced Arrhythmias
- PMID: 24962810
- PMCID: PMC4293372
- DOI: 10.1002/jcp.24709
Characterization of Cardiac Anoctamin1 Ca²⁺-Activated Chloride Channels and Functional Role in Ischemia-Induced Arrhythmias
Abstract
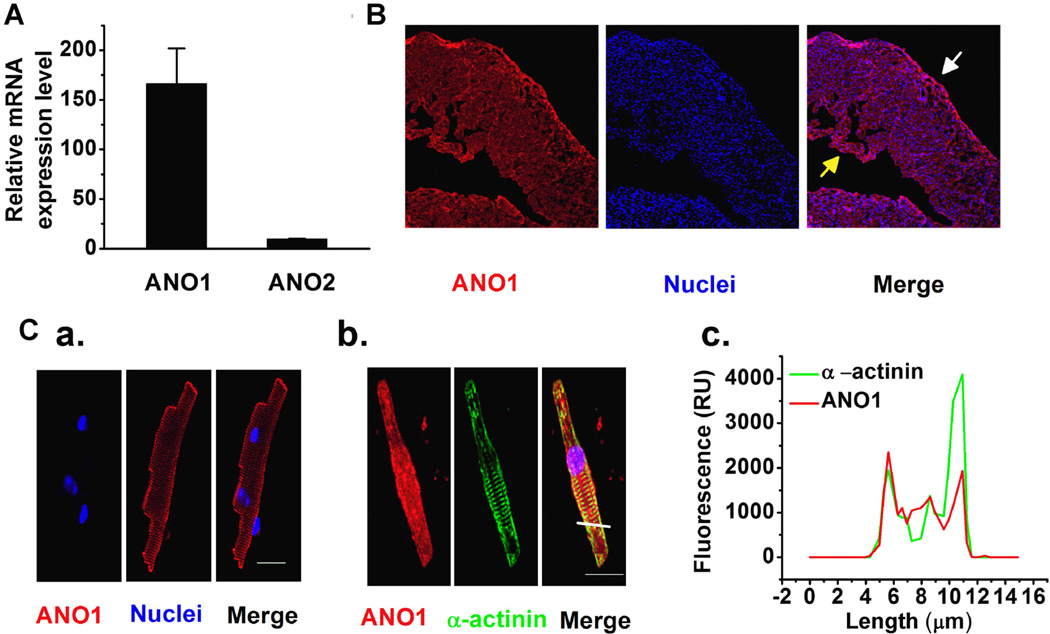
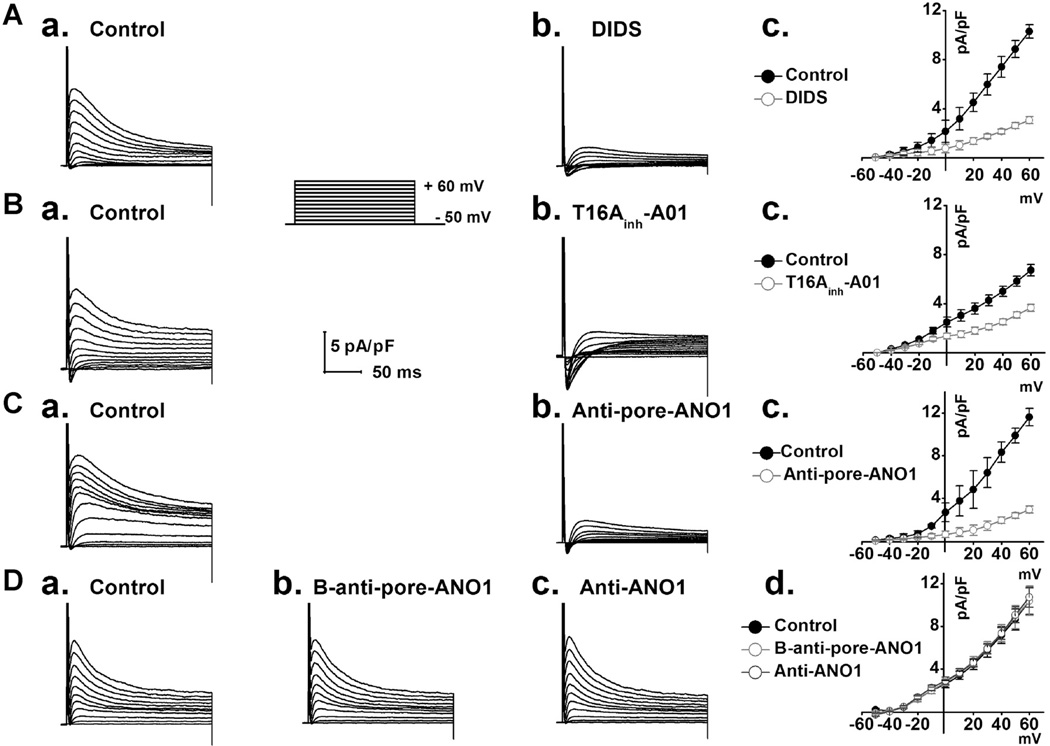
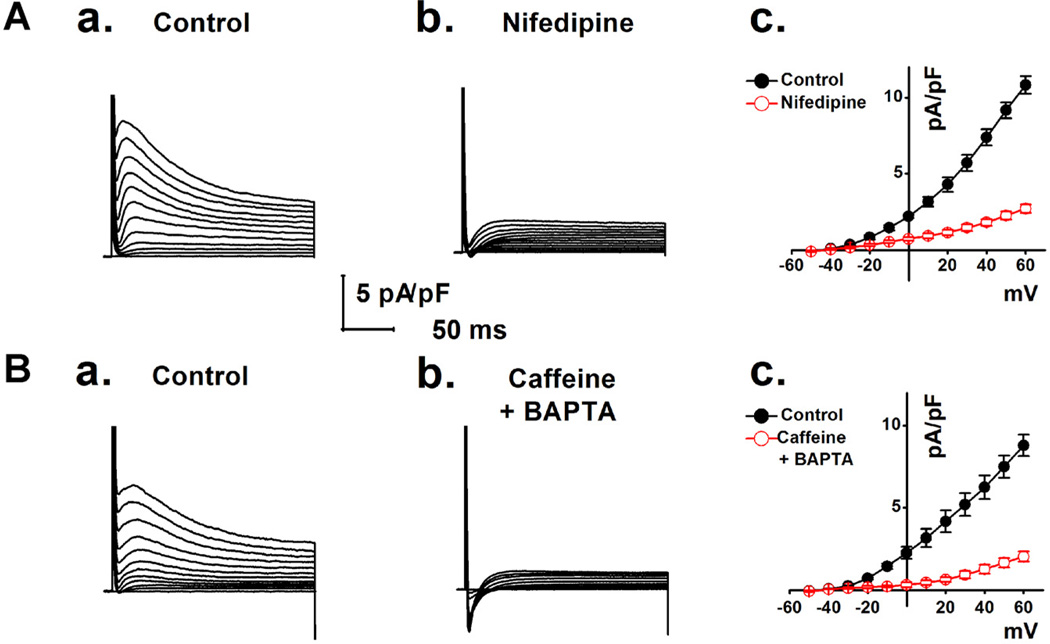
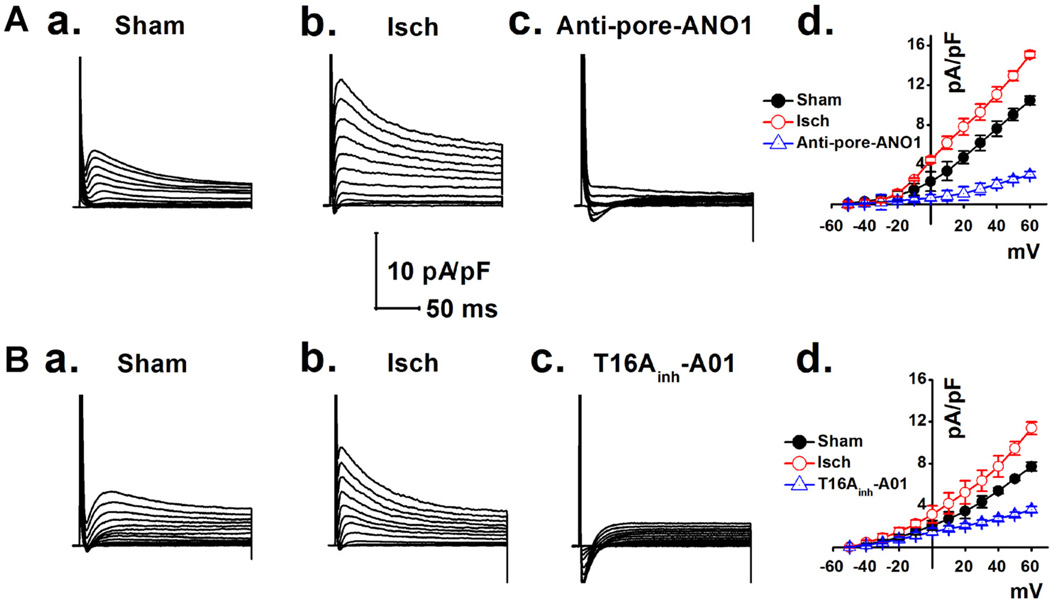
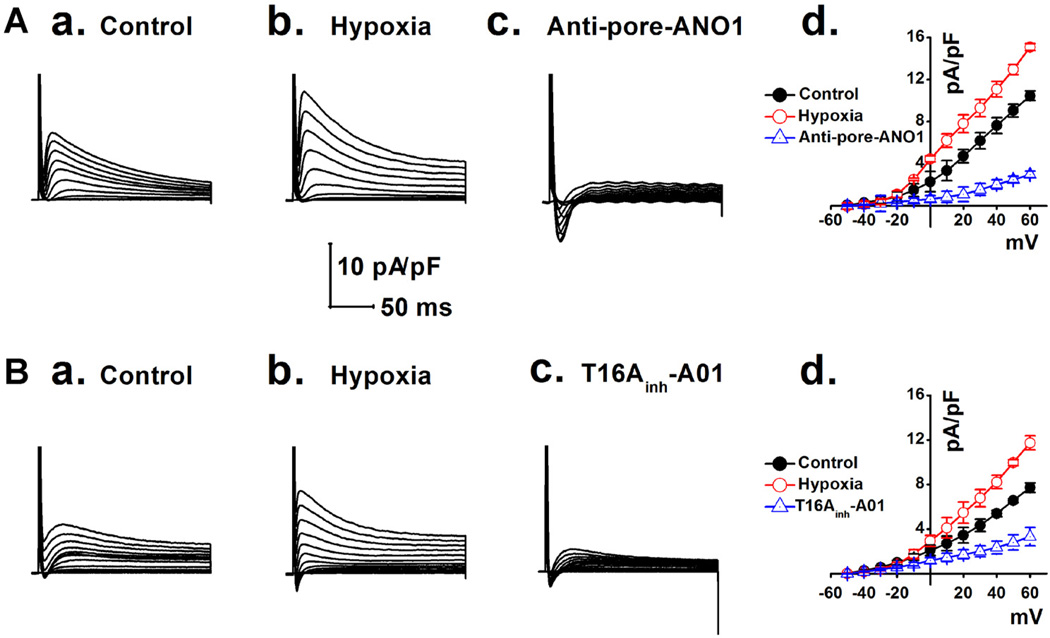
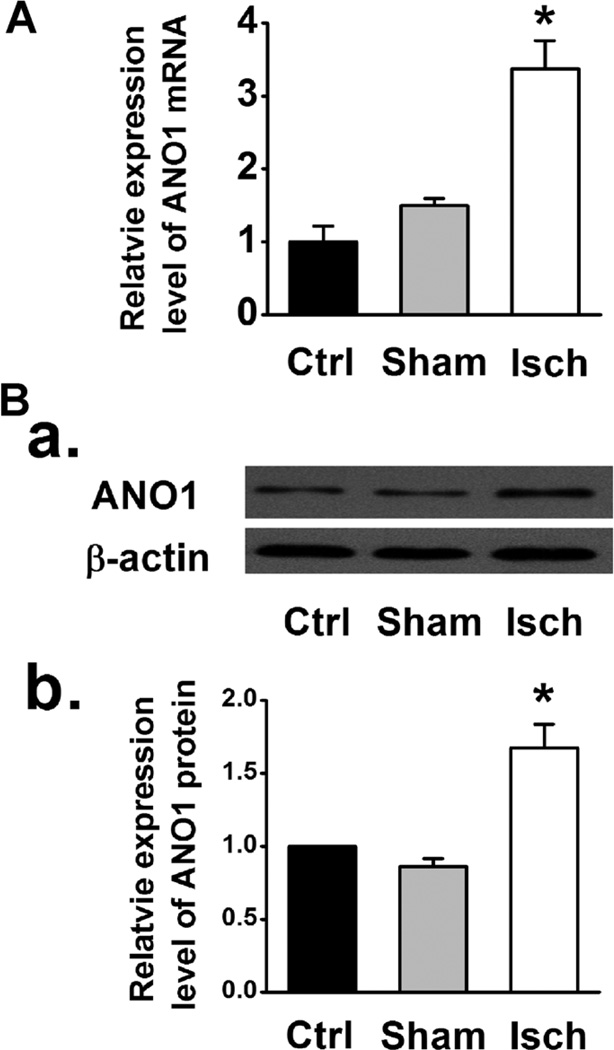
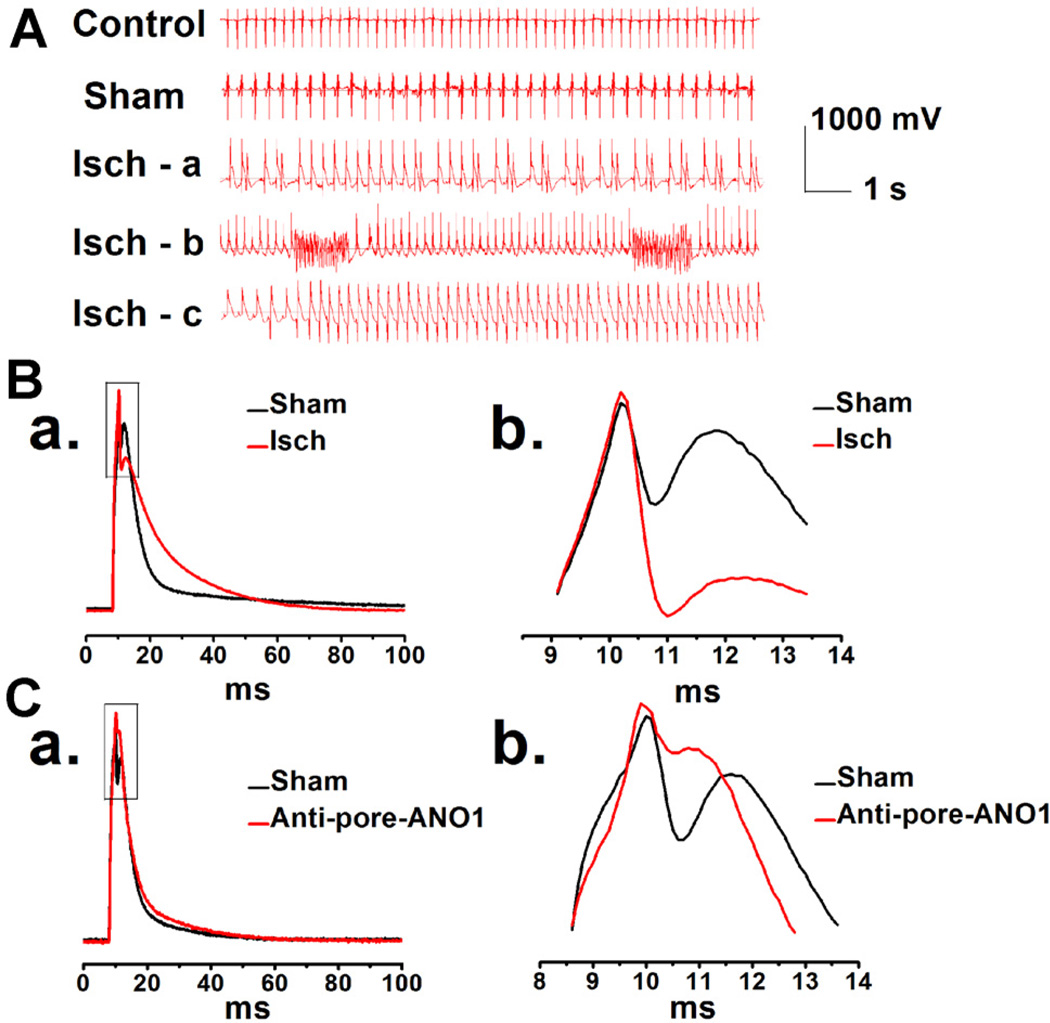
Anoctamin1 (ANO1) encodes a Ca(2+)-activated chloride (Cl(-)) channel (CaCC) in variety tissues of many species. Whether ANO1 expresses and functions as a CaCC in cardiomyocytes remain unknown. The objective of this study is to characterize the molecular and functional expression of ANO1 in cardiac myocytes and the role of ANO1-encoded CaCCs in ischemia-induced arrhythmias in the heart. Quantitative real-time RT-PCR, immunofluorescence staining assays, and immunohistochemistry identified the molecular expression, location, and distribution of ANO1 in mouse ventricular myocytes (mVMs). Patch-clamp recordings combined with pharmacological analyses found that ANO1 was responsible for a Ca(2+)-activated Cl(-) current (I(Cl.Ca)) in cardiomyocytes. Myocardial ischemia led to a significant increase in the current density of I(Cl.Ca), which was inhibited by a specific ANO1 inhibitor, T16A(inh)-A01, and an antibody targeting at the pore area of ANO1. Moreover, cardiomyocytes isolated from mice with ischemia-induced arrhythmias had an accelerated early phase 1 repolarization of action potentials (APs) and a deeper "spike and dome" compared to control cardiomyocytes from non-ischemia mice. Application of the antibody targeting at ANO1 pore prevented the ischemia-induced early phase 1 repolarization acceleration and caused a much shallower "spike and dome". We conclude that ANO1 encodes CaCC and plays a significant role in the phase 1 repolarization of APs in mVMs. The ischemia-induced increase in ANO1 expression may be responsible for the increased density of I(Cl.Ca) in the ischemic heart and may contribute, at least in part, to ischemia-induced arrhythmias.
© 2014 Wiley Periodicals, Inc.
Figures

Similar articles
-
Anoctamin-1 Cl(-) channels in nociception: activation by an N-aroylaminothiazole and capsaicin and inhibition by T16A[inh]-A01.Mol Pain. 2015 Sep 12;11:55. doi: 10.1186/s12990-015-0061-y. Mol Pain. 2015. PMID: 26364309 Free PMC article.
-
Coronary hypercontractility to acidosis owes to the greater activity of TMEM16A/ANO1 in the arterial smooth muscle cells.Biomed Pharmacother. 2021 Jul;139:111615. doi: 10.1016/j.biopha.2021.111615. Epub 2021 May 7. Biomed Pharmacother. 2021. PMID: 34243598
-
Hypoxia augments the calcium-activated chloride current carried by anoctamin-1 in cardiac vascular endothelial cells of neonatal mice.Br J Pharmacol. 2014 Aug;171(15):3680-92. doi: 10.1111/bph.12730. Br J Pharmacol. 2014. PMID: 24758567 Free PMC article.
-
Activation of Ca(2+) -activated Cl(-) channel ANO1 by localized Ca(2+) signals.J Physiol. 2016 Jan 1;594(1):19-30. doi: 10.1113/jphysiol.2014.275107. Epub 2014 Dec 15. J Physiol. 2016. PMID: 25398532 Free PMC article. Review.
-
Molecular mechanisms of activation and regulation of ANO1-Encoded Ca2+-Activated Cl- channels.Channels (Austin). 2021 Dec;15(1):569-603. doi: 10.1080/19336950.2021.1975411. Channels (Austin). 2021. PMID: 34488544 Free PMC article. Review.
Cited by
-
Electrophysiological and Molecular Mechanisms of Sinoatrial Node Mechanosensitivity.Front Cardiovasc Med. 2021 Aug 9;8:662410. doi: 10.3389/fcvm.2021.662410. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34434970 Free PMC article.
-
Ca2+-activated Cl- current is antiarrhythmic by reducing both spatial and temporal heterogeneity of cardiac repolarization.J Mol Cell Cardiol. 2017 Aug;109:27-37. doi: 10.1016/j.yjmcc.2017.06.014. Epub 2017 Jun 29. J Mol Cell Cardiol. 2017. PMID: 28668303 Free PMC article.
-
TMEM16A Plays an Insignificant Role in Myocardium Remodeling but May Promote Angiogenesis of Heart During Pressure-overload.Front Physiol. 2022 May 31;13:897619. doi: 10.3389/fphys.2022.897619. eCollection 2022. Front Physiol. 2022. PMID: 35711304 Free PMC article.
-
Calcium-activated chloride current determines action potential morphology during calcium alternans in atrial myocytes.J Physiol. 2016 Feb 1;594(3):699-714. doi: 10.1113/JP271887. Epub 2016 Jan 15. J Physiol. 2016. PMID: 26662365 Free PMC article.
-
Telocytes: Supporting cells participating in ventricular arrhythmogenesis?J Arrhythm. 2019 Apr 15;35(3):522-523. doi: 10.1002/joa3.12181. eCollection 2019 Jun. J Arrhythm. 2019. PMID: 31293702 Free PMC article. No abstract available.
References
-
- Bozeat ND, Xiang SY, Ye LL, Yao TY, Duan ML, Burkin DJ, Lamb FS, Duan DD. Activation of volume regulated chloride channels protects myocardium from ischemia/reperfusion damage in second-window ischemic preconditioning. Cell Physiol Biochem: Int J Exp Cell Physiol Biochem Pharmacol. 2011;28:1265–1278. - PMC - PubMed
-
- Caputo A, Caci E, Ferrera L, Pedemonte N, Barsanti C, Sondo E, Pfeffer U, Ravazzolo R, Zegarra-Moran O, Galietta LJ. TMEM 16A, a membrane protein associated with calcium-dependent chloride channel activity. Science. 2008;322:590–594. - PubMed
-
- Collier ML, Levesque PC, Kenyon JL, Hume JR. Unitary Cl− channels activated by cytoplasmic Ca2+ in canine ventricular myocytes. Circ Res. 1996;78:936–944. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
