

Review
doi: 10.1007/978-94-017-9050-5_7.
The roles of retinoic acid and retinoic acid receptors in inducing epigenetic changes
Affiliations
- PMID: 24962884
- PMCID: PMC4199334
- DOI: 10.1007/978-94-017-9050-5_7
Item in Clipboard
Review
The roles of retinoic acid and retinoic acid receptors in inducing epigenetic changes
Subcell Biochem.
2014.
Abstract
Epigenetics is "the branch of biology which studies the causal interactions between genes and their products which bring the phenotype into being" as defined by Conrad Waddington in 1942 in a discussion of the mechanisms of cell differentiation. More than seven decades later we know that these mechanisms include histone tail post-translational modifications, DNA methylation, ATP-dependent chromatin remodeling, and non-coding RNA pathways. Epigenetic modifications are powerful drugs targets, and combined targeting of multiple pathways is expected to significantly advance cancer therapy.
Figures
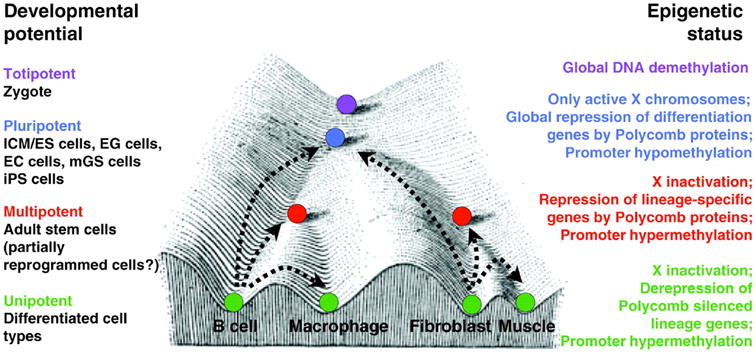
Conrad Waddington's Epigenetic Landscape illustrates the causal mechanisms by which the genotype brings about phenotypic effects. The progressive cellular differentiation (represented by purple, blue, red, and green) into specific cell types (illustrated by the different valleys) is associated with distinct epigenetic modifications. Polycomb repression and promoter hypermethylation are key mechanisms in the regulation of cellular differentiation. The corresponding epigenetic status of the various differentiation states is depicted on the right. (Hochedlinger and Plath, 2009).
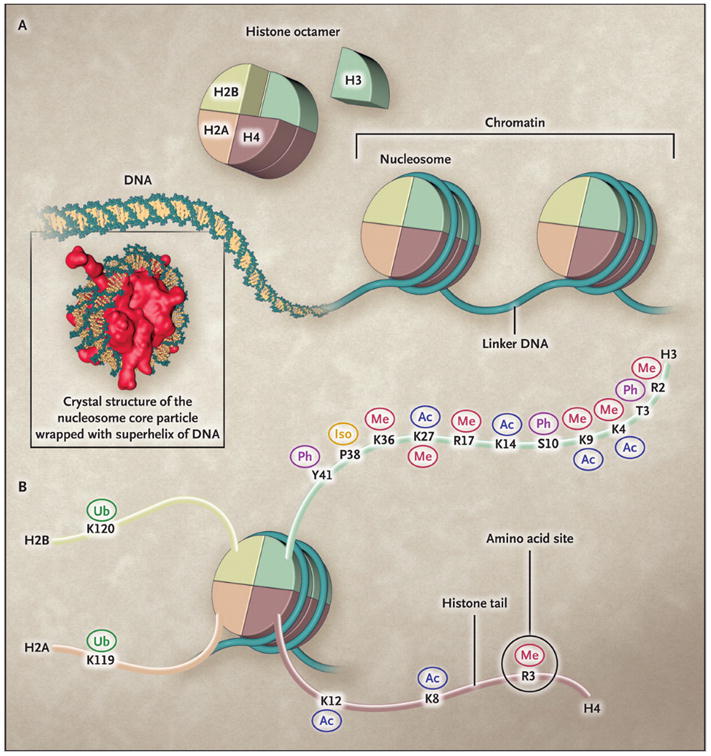
The basic functional unit of chromatin is the nucleosome (Panel A), which is composed of a histone octamer around which DNA is wrapped. Octamers are separated by linker DNA. The histone octamer is assembled from a histone H3:H4 tetramer and two H2A:H2B dimers. The histone tails of all four core histones are subject to a variety of post-translational modifications (Panel B). These include methylation (Me), acetylation (Ac), phosphorylation (Ph), ubiquitylation (Ub), and proline isomerization (Iso), all of which occur at the site of a specific amino acid, such as K4 and K9 on the histone H3 tail. The same histone amino acid may be subject to different post-translational modifications, which may facilitate different biologic outcomes. (Dawson et al., 2012).
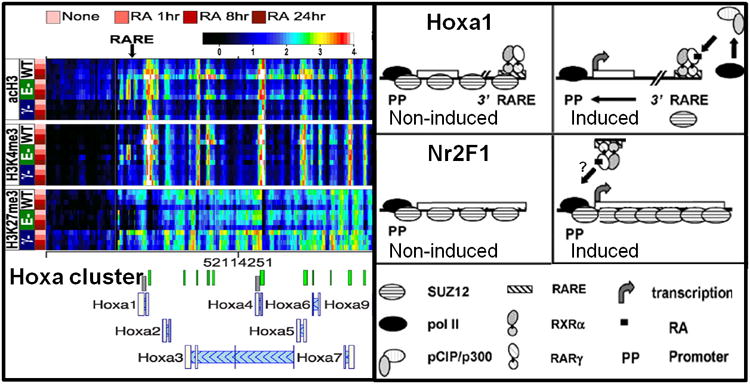
The epigenetic changes of the RA responsive Hoxa gene cluster are shown, with the locations of the Hoxa1 proximal promoter (PP) and RA responsive element (RARE) indicated by arrows. The levels of acH3, H3K4me3, and H3K27me3 determined by ChIP-chip are presented as heatmaps, with rows representing individual timepoints for each genotype, and columns indicating specific genomic regions. The genotypes of thestem cell lines are as follows: WT, RARE-KO (E-), and RARγ-KO (γ-). The cells were cultured in RA for 1, 8, and 24 h, as indicated. The color scale representing log2-transformed ChIP enrichment is indicated at the top of the figure. Note the reduced levels of H3ac and H3K4me3 at Hoxa1 PP and RARE in the RARγ-KO cell line Models for RA mediated transcription of RA target genes Hoxa1 and Nr2F1. In the absence of RA, RARγ-RXRαheterodimers associated with Hoxa1 RAREs presumably associate with co-repressors, thereby generating a SUZ12-rich environment which represses transcription. Binding of the RA ligand causes a conformational change in the RARγ-RXRα heterodimer bound to the Hoxa1 RARE. This results in the recruitment of pCIP/p300, which generates an euchromatic environment, presumably by acetylating the histone tails. This allows pol II to initiate transcription of Hoxa1. The Nr2F1 promoter region (PP) is bound by SUZ12 in the absence of RA. Upon exposure to RA the increase in activating marks is initially counteracted by a concomitant increase in SUZ12, which attenuates the transcription of Nr2F1. Eventually, the SUZ12 levels decline, allowing the increased transcriptional activation of Nr2F1. (modified from Kashyap et al., 2011 and Gillespie and Gudas, 2007b).
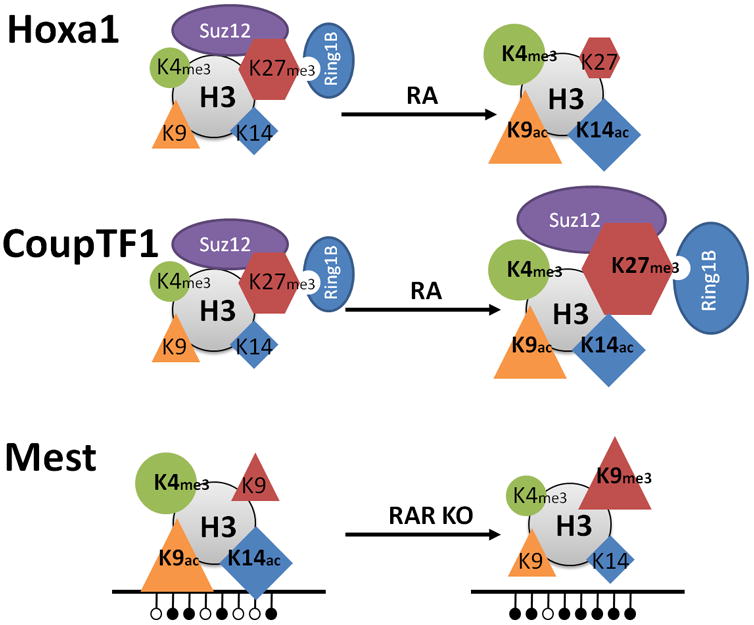
Hoxa1 represents a groupof direct target genes induced by RA (upper panel). The induction is characterized by dissociation of PRCs (ovals) and depletion of the H3K27me3 repressive mark (hexagons), and by increased levels of transcriptionally permissive marks, H3K4me3 (circle), H3K9ac (triangle), and H3K14ac (diamond). CoupTF1 representsa group of target genes with delayed transcriptional induction by RA (middle panel). The induction is characterized by an initial increase of PRCs (ovals) and of the H3K27me3 repressive mark (hexagons), concurrent with increased levels of transcriptionally permissive marks; H3K4me3, H3K9ac, and H3K14ac. The imprinted gene Mest is transcribed in the presence of RARα, but is silenced by DNA methylation upon knockout of RARα (lower panel). The transcriptional silencing of Mest is associated with increased DNA methylation, increased levels of the H3K9me3 repressive mark, and with decreased levels of transcriptionally permissive marks; H3K4me3, H3K9ac, and H3K14ac. Note that for Hoxa1 and CoupTF1 the transcriptionally active state is shown to the right, whereas for Mest the transcriptionally active state is shown to the left, Published in parts (CoupTF1) in Nucleic Acid Research (modified from Laursen, et al., 2013).
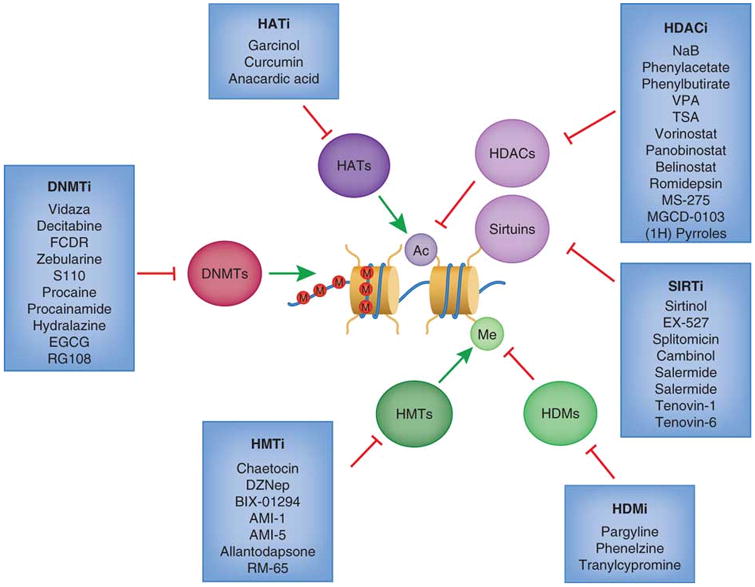
Histone acetyltransferase inhibitors (HATi) may be effective treatments since HAT proteins are sometimes considered to be tumor promoters. Histone deacetylase inhibitors (HDACi) such as vorinostat (SAHA) and romidepsin have been FDA approved for treatments of certain hematological cancers. SIRT inhibitors (SIRTi) specifically target the SIRT family of histone deacetylases. Histone demethyltransferase inhibitors (HDMi) and histone methylasetransferase (HMTi) have importantly become more selective towards specific marks, since methylation can act as both a repressive and an activating mark. DNA methyltransferase inhibitors (DNMTi) may serve as promising drugs for re-sensitizing certain cancer cells to chemotherapy, and two (vidaza and decitabine) are FDA approved for cancer treatment. (Rodríguez-Paredes et al., 2011)
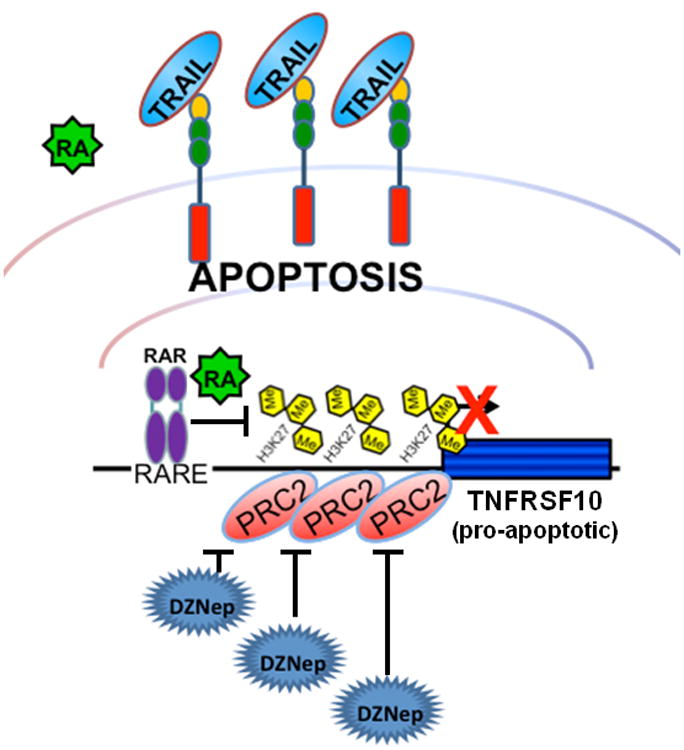
Retinoic acid treatment promotes TRAIL-related apoptosis in RARβ/RARγ–positive HT29 cells, but not in SW480 cells which express only low levels of RARβ/RARγ. The functional depletion of PRC2 by either inhibition with DZNep or by knockdown of SUZ12 increases TRAIL-mediated apoptosis in both HT29 and SW480 cell lines. In this scenario the PRC2-mediated repression is alleviated, thereby activating TNFRSF10 even in the absence of RARβ/RARγ.
Similar articles
-
Signals and combinatorial functions of histone modifications.Annu Rev Biochem. 2011;80:473-99. doi: 10.1146/annurev-biochem-061809-175347. Annu Rev Biochem. 2011. PMID: 21529160 Review.
-
Three-dimensional regulation of transcription.Protein Cell. 2015 Apr;6(4):241-53. doi: 10.1007/s13238-015-0135-7. Epub 2015 Feb 12. Protein Cell. 2015. PMID: 25670626 Free PMC article. Review.
-
Epigenetic regulation in human melanoma: past and future.Epigenetics. 2015;10(2):103-21. doi: 10.1080/15592294.2014.1003746. Epigenetics. 2015. PMID: 25587943 Free PMC article. Review.
-
Epigenetics: judge, jury and executioner of stem cell fate.Epigenetics. 2012 Aug;7(8):823-40. doi: 10.4161/epi.21141. Epub 2012 Jul 18. Epigenetics. 2012. PMID: 22805743 Free PMC article. Review.
-
Decoding liver injury: A regulatory role for histone modifications.Int J Biochem Cell Biol. 2015 Oct;67:188-93. doi: 10.1016/j.biocel.2015.03.009. Epub 2015 Mar 20. Int J Biochem Cell Biol. 2015. PMID: 25801055 Review.
Cited by
-
Epigenetic Cancer Therapy.Indian J Surg Oncol. 2024 Sep;15(3):447-450. doi: 10.1007/s13193-024-02067-w. Epub 2024 Aug 24. Indian J Surg Oncol. 2024. PMID: 39239429 Free PMC article. No abstract available.
-
Control of Innate and Adaptive Lymphocytes by the RAR-Retinoic Acid Axis.Immune Netw. 2018 Jan 18;18(1):e1. doi: 10.4110/in.2018.18.e1. eCollection 2018 Feb. Immune Netw. 2018. PMID: 29503736 Free PMC article. Review.
-
Role of retinoids in the prevention and treatment of colorectal cancer.World J Gastrointest Oncol. 2015 Oct 15;7(10):184-203. doi: 10.4251/wjgo.v7.i10.184. World J Gastrointest Oncol. 2015. PMID: 26483874 Free PMC article. Review.
-
The Mitochondrial Protein VDAC1 at the Crossroads of Cancer Cell Metabolism: The Epigenetic Link.Cancers (Basel). 2020 Apr 22;12(4):1031. doi: 10.3390/cancers12041031. Cancers (Basel). 2020. PMID: 32331482 Free PMC article.
-
Identification of Ethanol and 4-Nitroquinoline-1-Oxide Induced Epigenetic and Oxidative Stress Markers During Oral Cavity Carcinogenesis.Alcohol Clin Exp Res. 2015 Aug;39(8):1360-72. doi: 10.1111/acer.12772. Alcohol Clin Exp Res. 2015. PMID: 26207766 Free PMC article.
References
-
- Agger K, Cloos PA, Christensen J, Pasini D, Rose S, Rappsilber J, Issaeva I, Canaani E, Salcini AE, Helin K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature. 2007;449:731–734. - PubMed
-
- Bantscheff M, Hopf C, Savitski MM, Dittmann A, Grandi P, Michon AM, Schlegl J, Abraham Y, Becher I, Bergamini G, Boesche M, Delling M, Dumpelfeld B, Eberhard D, Huthmacher C, Mathieson T, Poeckel D, Reader V, Strunk K, Sweetman G, Kruse U, Neubauer G, Ramsden NG, Drewes G. Chemoproteomics profiling of HDAC inhibitors reveals selective targeting of HDAC complexes. Nat Biotechnol. 2011;29:255–265. - PubMed
-
- Béguelin W, Popovic R, Teater M, Jiang Y, Bunting KL, Rosen M, Shen H, Yang SN, Wang L, Ezponda T, Martinez-Garcia E, Zhang H, Zheng Y, Verma SK, McCabe MT, Ott HM, Van Aller GS, Kruger RG, Liu Y, McHugh CF, Scott DW, Chung YR, Kelleher N, Shaknovich R, Creasy CL, Gascoyne RD, Wong KK, Cerchietti L, Levine RL, Abdel-Wahab O, Licht JD, Elemento O, Melnick AM. EZH2 Is Required for Germinal Center Formation and Somatic EZH2 Mutations Promote Lymphoid Transformation. Cancer Cell. 2013;23:677–692. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
