

The evolution of parasitism in Nematoda
- PMID: 24963797
- PMCID: PMC4413787
- DOI: 10.1017/S0031182014000791
The evolution of parasitism in Nematoda
Abstract
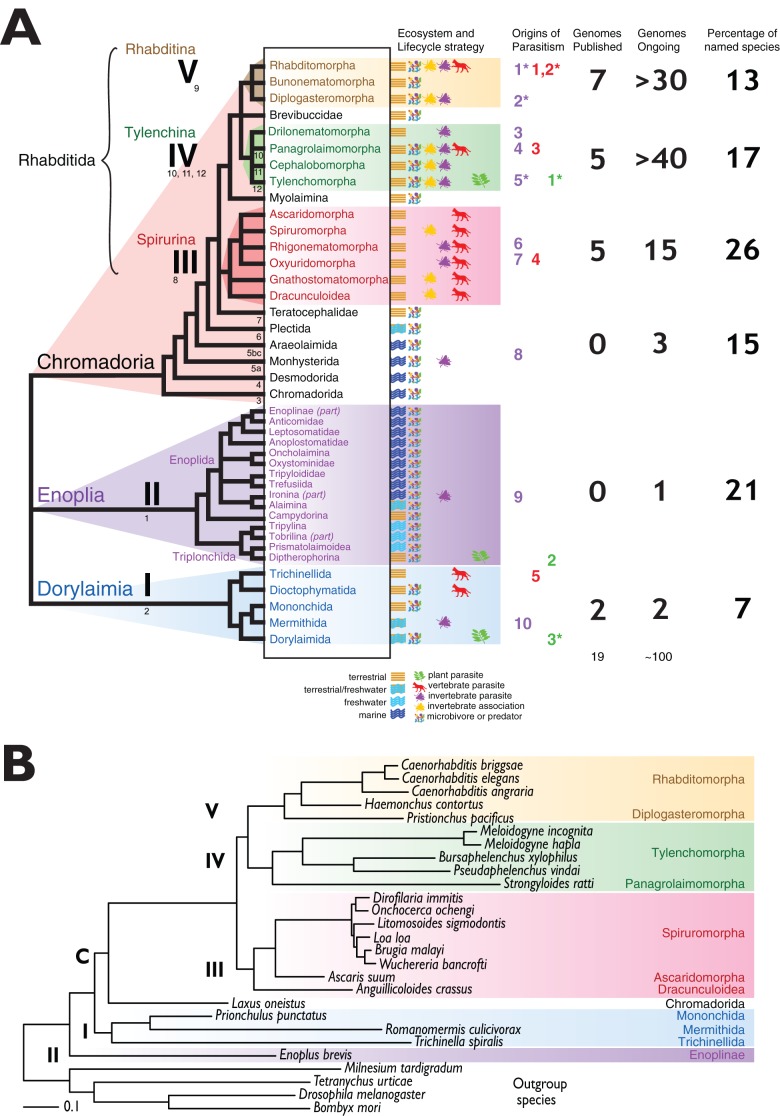
Nematodes are abundant and diverse, and include many parasitic species. Molecular phylogenetic analyses have shown that parasitism of plants and animals has arisen at least 15 times independently. Extant nematode species also display lifestyles that are proposed to be on the evolutionary trajectory to parasitism. Recent advances have permitted the determination of the genomes and transcriptomes of many nematode species. These new data can be used to further resolve the phylogeny of Nematoda, and identify possible genetic patterns associated with parasitism. Plant-parasitic nematode genomes show evidence of horizontal gene transfer from other members of the rhizosphere, and these genes play important roles in the parasite-host interface. Similar horizontal transfer is not evident in animal parasitic groups. Many nematodes have bacterial symbionts that can be essential for survival. Horizontal transfer from symbionts to the nematode is also common, but its biological importance is unclear. Over 100 nematode species are currently targeted for sequencing, and these data will yield important insights into the biology and evolutionary history of parasitism. It is important that these new technologies are also applied to free-living taxa, so that the pre-parasitic ground state can be inferred, and the novelties associated with parasitism isolated.
Keywords: horizontal gene transfer.
Figures
References
-
- Abad P., Gouzy J., Aury J. M., Castagnone-Sereno P., Danchin E. G., Deleury E., Perfus-Barbeoch L., Anthouard V., Artiguenave F., Blok V. C., Caillaud M. C., Coutinho P. M., Dasilva C., De Luca F., Deau F., Esquibet M., Flutre T., Goldstone J. V., Hamamouch N., Hewezi T., Jaillon O., Jubin C., Leonetti P., Magliano M., Maier T. R., Markov G. V., McVeigh P., Pesole G., Poulain J., Robinson-Rechavi M., Sallet E., Segurens B., Steinbach D., Tytgat T., Ugarte E., van Ghelder C., Veronico P., Baum T. J., Blaxter M., Bleve-Zacheo T., Davis E. L., Ewbank J. J., Favery B., Grenier E., Henrissat B., Jones J. T., Laudet V., Maule A. G., Quesneville H., Rosso M. N., Schiex T., Smant G., Weissenbach J. and Wincker P. (2008). Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nature Biotechnology 26, 882–884. - PubMed
-
- Aguinaldo A. M. A., Turbeville J. M., Linford L. S., Rivera M. C., Garey J. R., Raff R. A. and Lake J. A. (1997). Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387, 489–493. - PubMed
-
- Bai X., Adams B. J., Ciche T. A., Clifton S., Gaugler R., Kim K. S., Spieth J., Sternberg P. W., Wilson R. K. and Grewal P. S. (2013). A lover and a fighter: the genome sequence of an entomopathogenic nematode Heterorhabditis bacteriophora. PLOS ONE 8, . doi: 10.1371/journal.pone.0069618. - DOI - PMC - PubMed
-
- Bandi C., McCall J. W., Genchi C., Corona S., Venco L. and Sacchi L. (1999). Effects of tetracycline on the filarial worms Brugia pahangi and Dirofilaria immitis and their bacterial endosymbionts Wolbachia. International Journal for Parasitology 29, 357–364. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
