

Immunogenicity of a synthetic vaccine based on Plasmodium vivax Duffy binding protein region II
- PMID: 24964808
- PMCID: PMC4178564
- DOI: 10.1128/CVI.00205-14
Immunogenicity of a synthetic vaccine based on Plasmodium vivax Duffy binding protein region II
Abstract
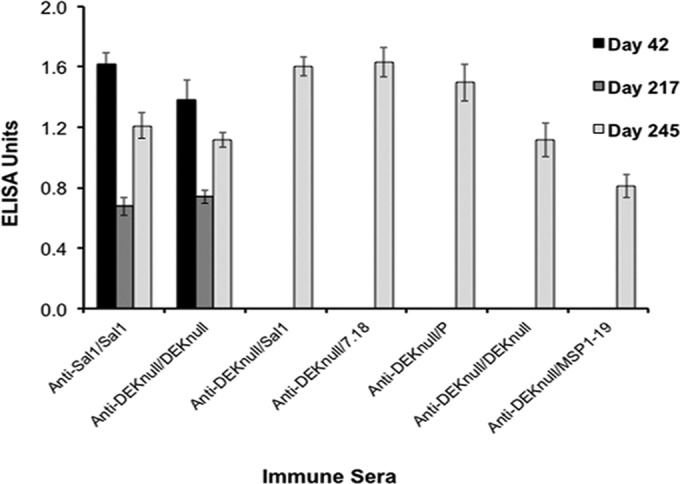
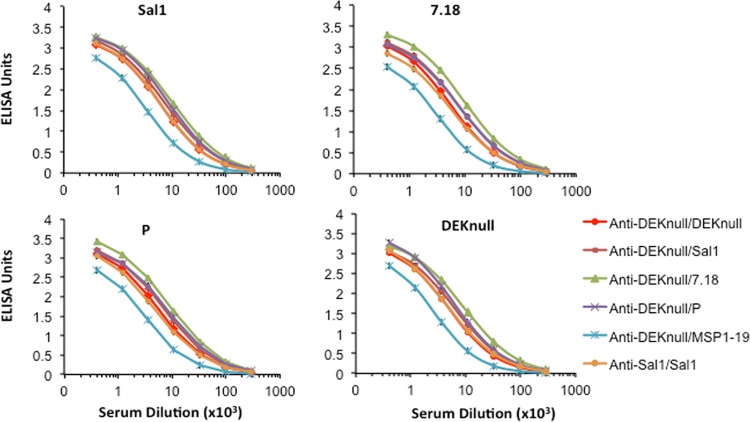
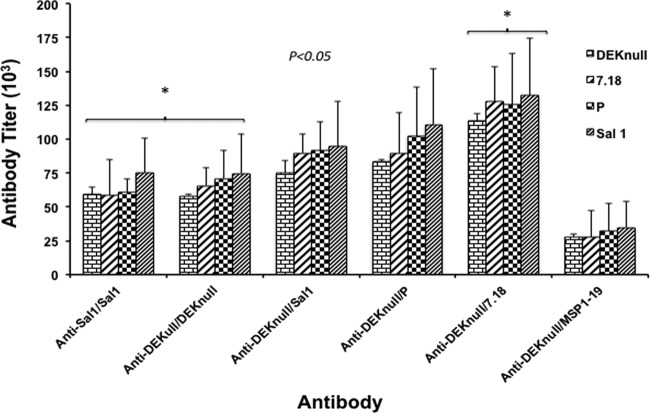
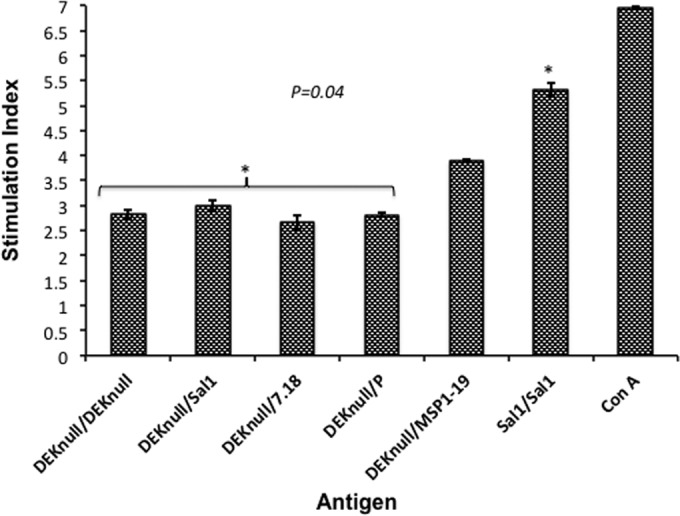
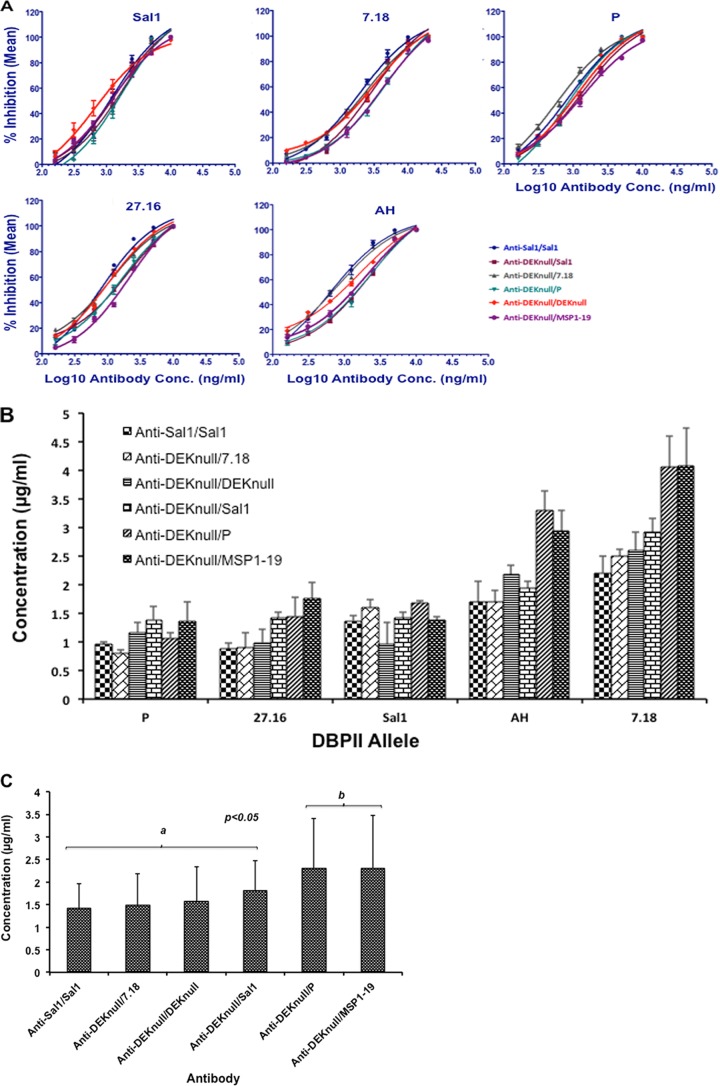
Molecules that play a role in Plasmodium merozoite invasion of host red blood cells represent attractive targets for blood-stage vaccine development against malaria. In Plasmodium vivax, merozoite invasion of reticulocytes is mediated by the Duffy binding protein (DBP), which interacts with its cognate receptor, the Duffy antigen receptor for chemokines, on the surface of reticulocytes. The DBP ligand domain, known as region II (DBPII), contains the critical residues for receptor recognition, making it a prime target for vaccine development against blood-stage vivax malaria. In natural infections, DBP is weakly immunogenic and DBPII allelic variation is associated with strain-specific immunity, which may compromise vaccine efficacy. In a previous study, a synthetic vaccine termed DEKnull that lacked an immunodominant variant epitope in DBPII induced functional antibodies to shared neutralizing epitopes on the native Sal1 allele. Anti-DEKnull antibody titers were lower than anti-Sal1 titers but produced more consistent, strain-transcending anti-DBPII inhibitory responses. In this study, we further characterized the immunogenicity of DEKnull, finding that immunization with recombinant DEKnull produced an immune response comparable to that obtained with native recombinant DBP alleles. Further investigation of DEKnull is necessary to enhance its immunogenicity and broaden its specificity.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Branch OH, Udhayakumar V, Hightower AW, Oloo AJ, Hawley WA, Nahlen BL, Bloland PB, Kaslow DC, Lal AA. 1998. A longitudinal investigation of IgG and IgM antibody responses to the merozoite surface protein-1 19-kiloDalton domain of Plasmodium falciparum in pregnant women and infants: associations with febrile illness, parasitemia, and anemia. Am. J. Trop. Med. Hyg. 58:211–219 - PubMed
-
- King CL, Michon P, Shakri AR, Marcotty A, Stanisic D, Zimmerman PA, Cole-Tobian JL, Mueller I, Chitnis CE. 2008. Naturally acquired Duffy-binding protein-specific binding inhibitory antibodies confer protection from blood-stage Plasmodium vivax infection. Proc. Natl. Acad. Sci. U. S. A. 105:8363–8368. 10.1073/pnas.0800371105 - DOI - PMC - PubMed
-
- Grimberg BT, Udomsangpetch R, Xainli J, McHenry A, Panichakul T, Sattabongkot J, Cui L, Bockarie M, Chitnis C, Adams J, Zimmerman PA, King CL. 2007. Plasmodium vivax invasion of human erythrocytes inhibited by antibodies directed against the Duffy binding protein. PLoS Med. 4(12):e337. 10.1371/journal.pmed.0040337 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
