

A novel partitivirus that confers hypovirulence on plant pathogenic fungi
- PMID: 24965462
- PMCID: PMC4136314
- DOI: 10.1128/JVI.01036-14
A novel partitivirus that confers hypovirulence on plant pathogenic fungi
Abstract
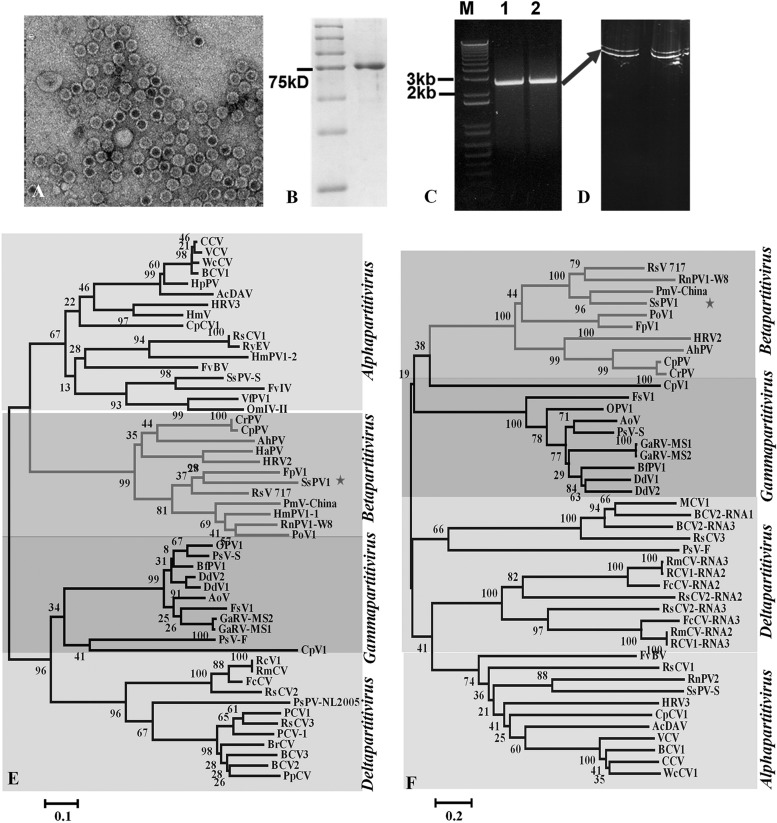
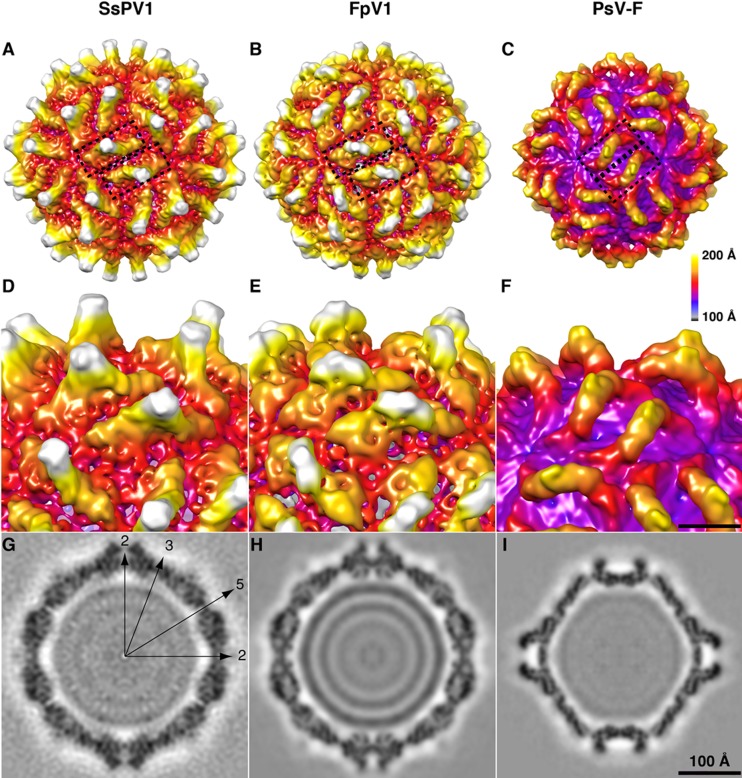
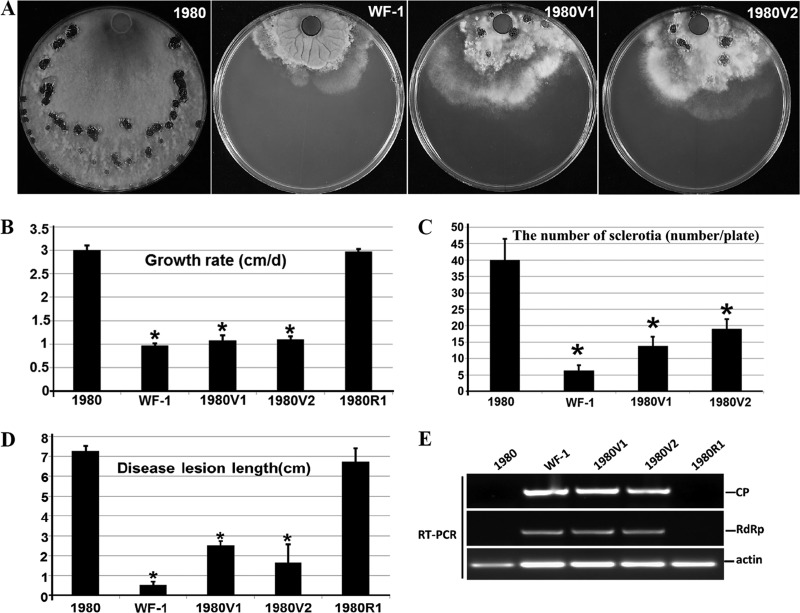
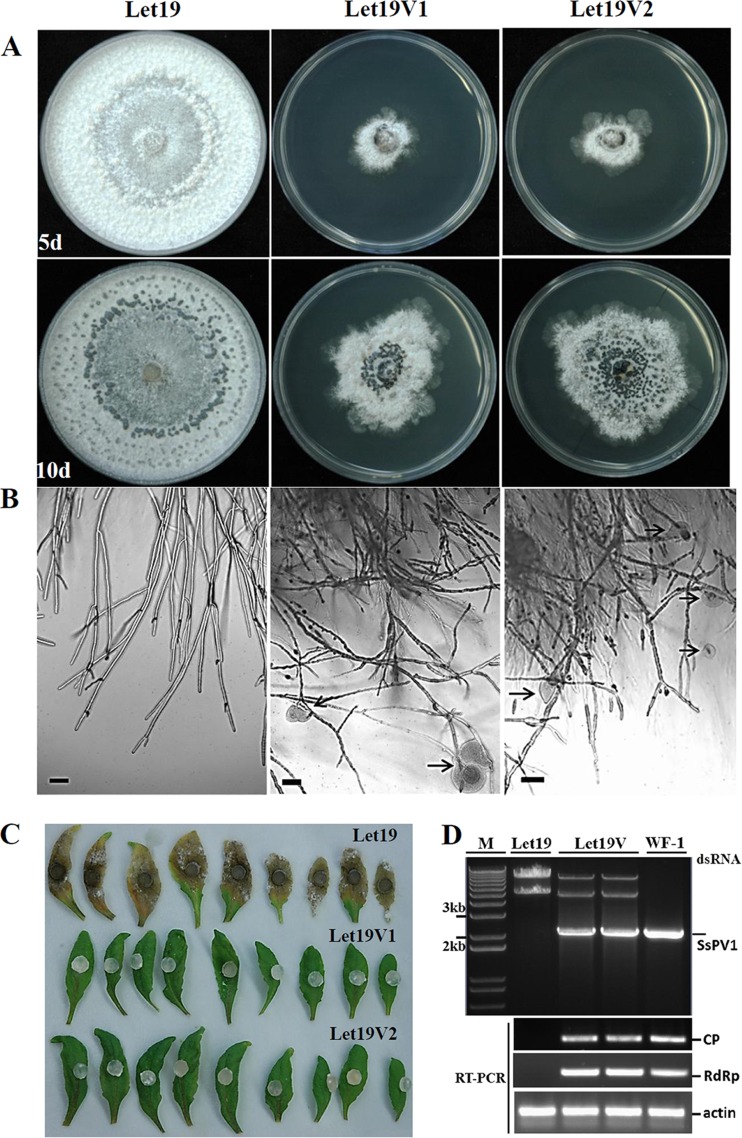
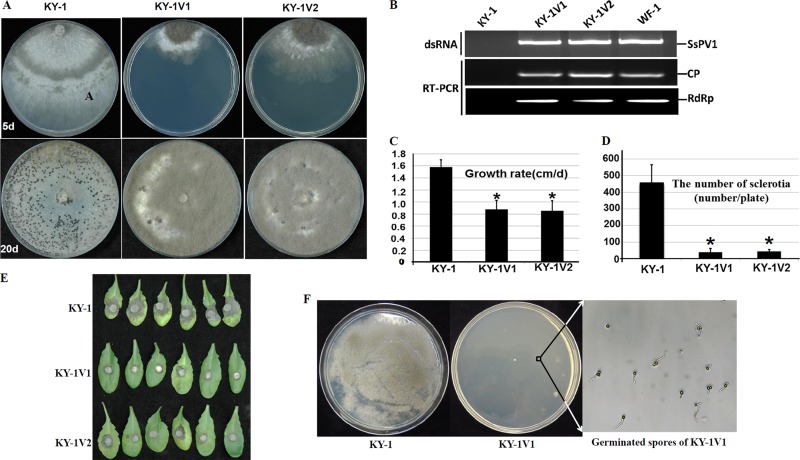
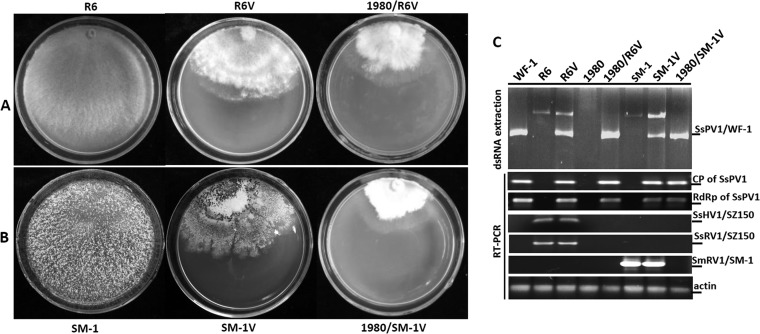
Members of the family Partitiviridae have bisegmented double-stranded RNA (dsRNA) genomes and are not generally known to cause obvious symptoms in their natural hosts. An unusual partitivirus, Sclerotinia sclerotiorum partitivirus 1 (SsPV1/WF-1), conferred hypovirulence on its natural plant-pathogenic fungal host, Sclerotinia sclerotiorum strain WF-1. Cellular organelles, including mitochondria, were severely damaged. Hypovirulence and associated traits of strain WF-1 and SsPV1/WF-1 were readily cotransmitted horizontally via hyphal contact to different vegetative compatibility groups of S. sclerotiorum and interspecifically to Sclerotinia nivalis and Sclerotinia minor. S. sclerotiorum strain 1980 transfected with purified SsPV1/WF-1 virions also exhibited hypovirulence and associated traits similar to those of strain WF-1. Moreover, introduction of purified SsPV1/WF-1 virions into strain KY-1 of Botrytis cinerea also resulted in reductions in virulence and mycelial growth and, unexpectedly, enhanced conidial production. However, virus infection suppressed hyphal growth of most germinating conidia of B. cinerea and was eventually lethal to infected hyphae, since very few new colonies could develop following germ tube formation. Taken together, our results support the conclusion that SsPV1/WF-1 causes hypovirulence in Sclerotinia spp. and B. cinerea. Cryo-EM (cryo-electron microscopy) reconstruction of the SsPV1 particle shows that it has a distinct structure with similarity to the closely related partitiviruses Fusarium poae virus 1 and Penicillium stoloniferum virus F. These findings provide new insights into partitivirus biological activities and clues about molecular interactions between partitiviruses and their hosts.
Importance: Members of the Partitiviridae are believed to occur commonly in their phytopathogenic fungal and plant hosts. However, most partitiviruses examined so far appear to be associated with latent infections. Here we report a partitivirus, SsPV1/WF-1, that was isolated from a hypovirulent strain of Sclerotinia sclerotiorum and describe its biological and molecular features. We have demonstrated that SsPV1 confers hypovirulence. Furthermore, SsPV1 can infect and cause hypovirulence in Botrytis cinerea. Our study also suggests that SsPV1 has a vigorous ability to proliferate and spread via hyphal contact. SsPV1 can overcome vegetative incompatibility barriers and can be transmitted horizontally among different vegetative compatibility groups of S. sclerotiorum, even interspecifically. Cryo-EM reconstruction of SsPV1 shows that it has a distinct structure with similarity to closely related partitiviruses. Our studies exploit a novel system, SsPV1 and its hosts, which can provide the means to explore the mechanisms by which partitiviruses interact with their hosts.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
