

Fine mapping of murine antibody responses to immunization with a novel soluble form of hepatitis C virus envelope glycoprotein complex
- PMID: 24965471
- PMCID: PMC4178869
- DOI: 10.1128/JVI.01584-14
Fine mapping of murine antibody responses to immunization with a novel soluble form of hepatitis C virus envelope glycoprotein complex
Erratum in
- J Virol. 2014 Nov;88(22):13517
Abstract
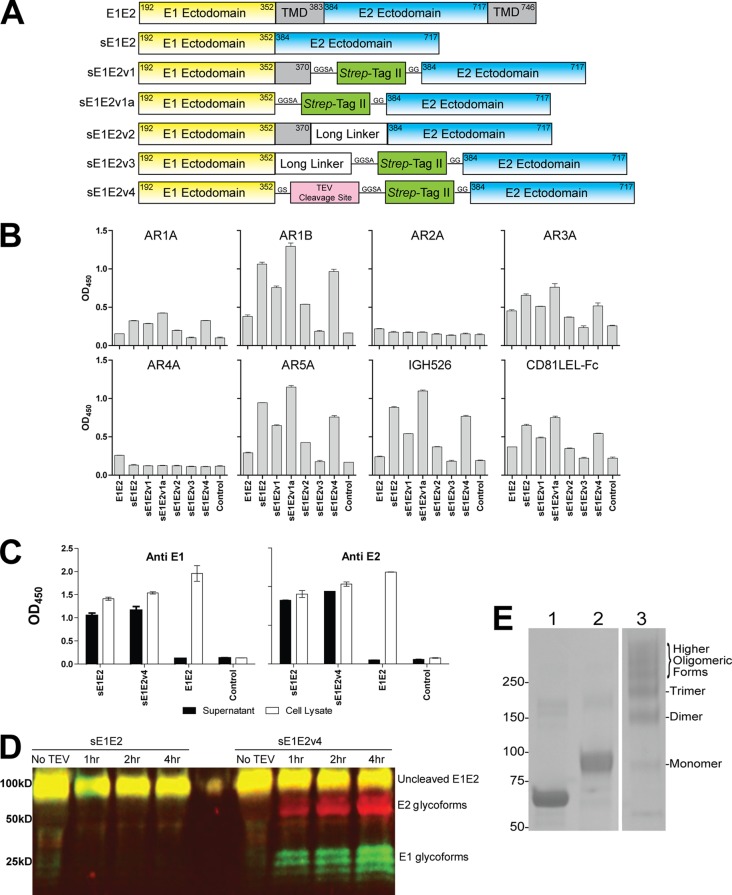
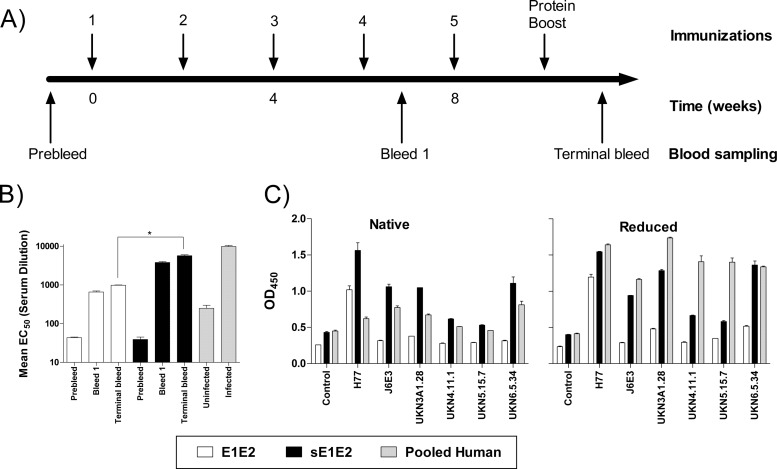
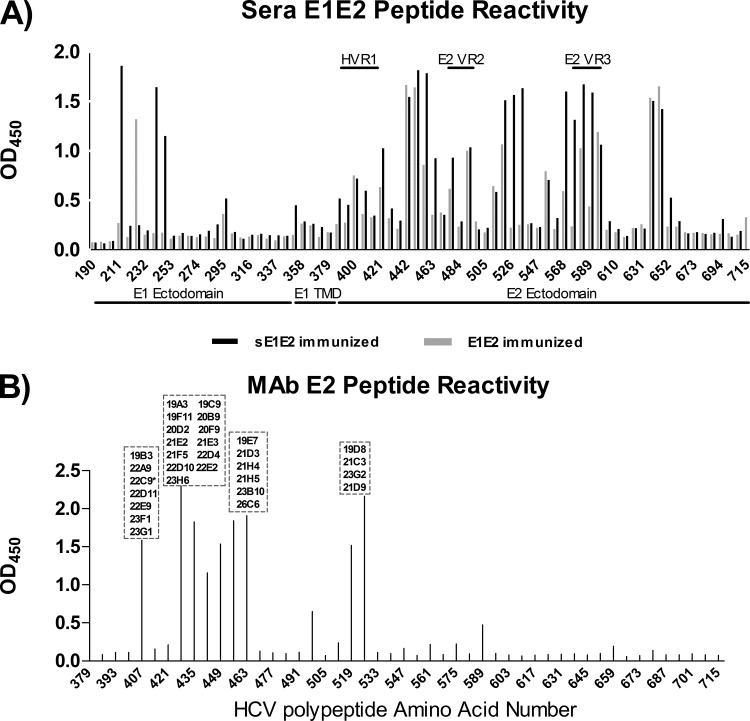
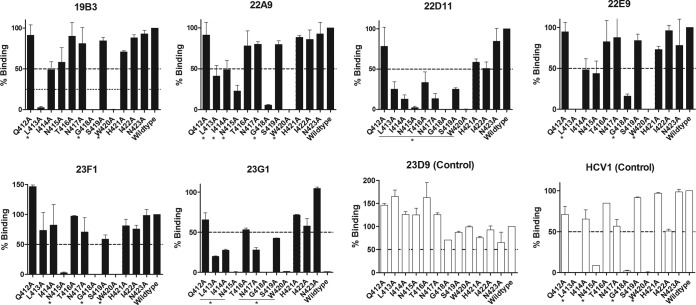
The hepatitis C virus (HCV) envelope glycoprotein E1E2 complex is a candidate vaccine antigen. Previous immunization studies of E1E2 have yielded various results on its ability to induce virus-neutralizing antibodies in animal models and humans. The murine model has become a vital tool for HCV research owing to the development of humanized mice susceptible to HCV infection. In this study, we investigated the antibody responses of mice immunized with E1E2 and a novel soluble form of E1E2 (sE1E2) by a DNA prime and protein boost strategy. The results showed that sE1E2 elicited higher antibody titers and a greater breadth of reactivity than the wild-type cell-associated E1E2. However, immune sera elicited by either immunogen were only weakly neutralizing. In order to understand the contrasting results of binding and serum neutralizing activities, epitopes targeted by the polyclonal antibody responses were mapped and monoclonal antibodies (MAbs) were generated. The results showed that the majority of serum antibodies were directed to the E1 region 211 to 250 and the E2 regions 421 to 469, 512 to 539, 568 to 609, and 638 to 651, instead of the well-known immunodominant E2 hypervariable region 1 (HVR1). Unexpectedly, in MAb analysis, ∼ 12% of MAbs isolated were specific to the conserved E2 antigenic site 412 to 423, and 85% of them cross-neutralized multiple HCV isolates. The epitopes recognized by these MAbs are similar but distinct from the previously reported HCV1 and AP33 broadly neutralizing epitopes. In conclusion, E1E2 can prime B cells specific to conserved neutralizing epitopes, but the levels of serum neutralizing antibodies elicited are insufficient for effective virus neutralization. The sE1E2 constructs described in this study can be a useful template for rational antigen engineering.
Importance: Hepatitis C virus infects 2 to 3% of the world's population and is a leading cause of liver failures and the need for liver transplantation. The virus envelope glycoprotein complex E1E2 produced by detergent extraction of cells overexpressing the protein was evaluated in a phase I clinical trial but failed to induce neutralizing antibodies in most subjects. In this study, we designed a novel form of E1E2 which is secreted from cells and is soluble and compared it to wild-type E1E2 by DNA immunization of mice. The results showed that this new E1E2 is more immunogenic than wild-type E1E2. Detailed mapping of the antibody responses revealed that antibodies to the conserved E2 antigenic site 412 to 423 were elicited but the serum concentrations were too low to neutralize the virus effectively. This soluble E1E2 provides a new reagent for studying HCV and for rational vaccine design.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Wasley A, Miller JT, Finelli L. 2007. Surveillance for acute viral hepatitis—United States, 2005. MMWR Surveill. Summ. 56(SS-03):1–24 - PubMed
-
- Leuchner L, Lindstrom H, Burstein GR, Mulhern KE, Rocchio EM, Johnson G, Schaffzin J, Smith P. 2008. Use of enhanced surveillance for hepatitis C virus infection to detect a cluster among young injection-drug users—New York, November 2004-April 2007. (Reprinted from MMWR 57:517–521, 2008). JAMA 300:34–36. 10.1001/jama.300.1.34 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
