

Modelling and mutational analysis of Aspergillus nidulans UreA, a member of the subfamily of urea/H⁺ transporters in fungi and plants
- PMID: 24966243
- PMCID: PMC4077062
- DOI: 10.1098/rsob.140070
Modelling and mutational analysis of Aspergillus nidulans UreA, a member of the subfamily of urea/H⁺ transporters in fungi and plants
Abstract
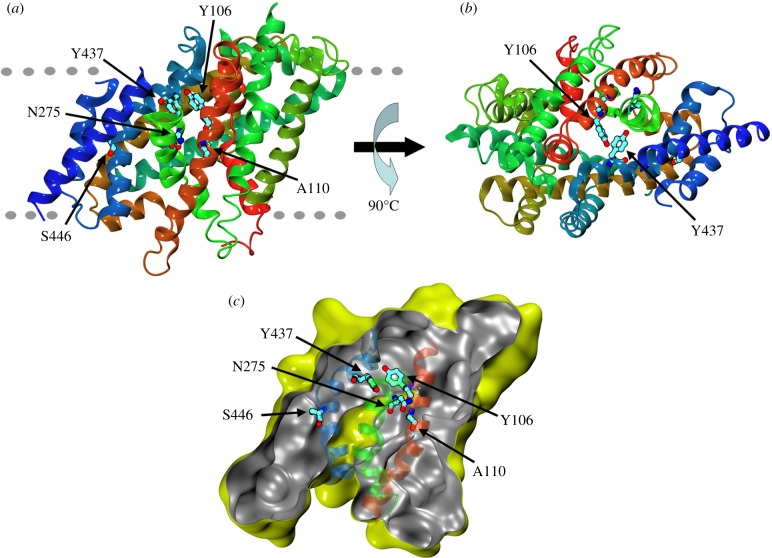
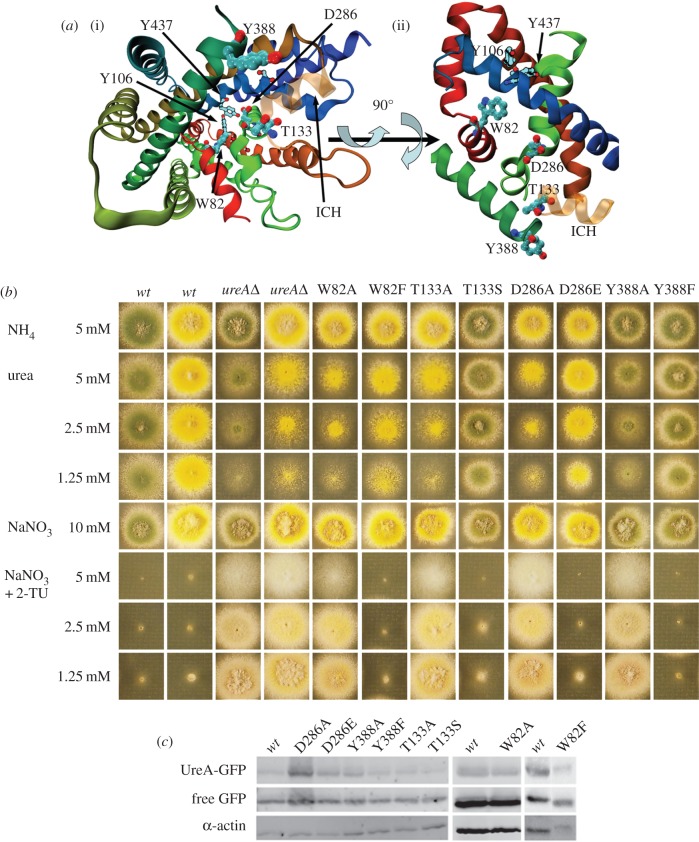
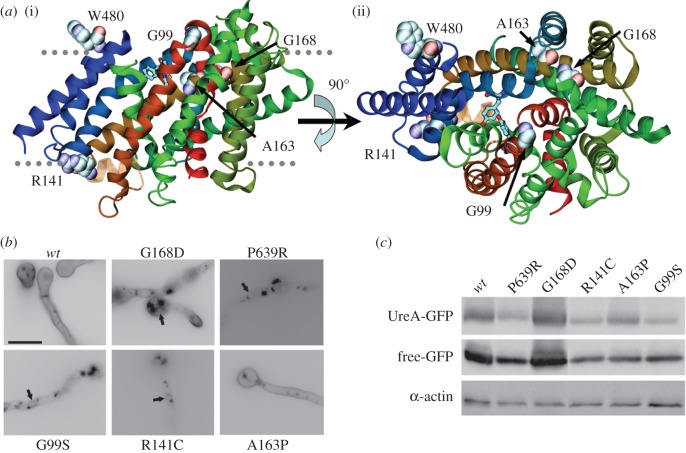
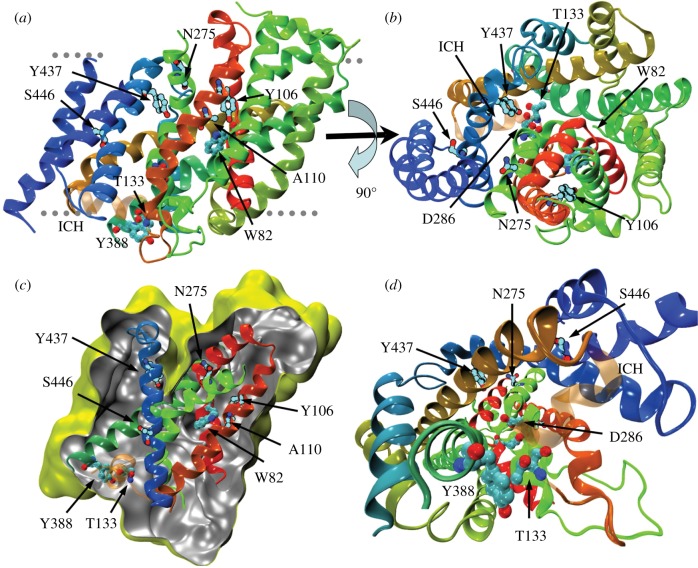
We present the first account of the structure-function relationships of a protein of the subfamily of urea/H(+) membrane transporters of fungi and plants, using Aspergillus nidulans UreA as a study model. Based on the crystal structures of the Vibrio parahaemolyticus sodium/galactose symporter (vSGLT) and of the Nucleobase-Cation-Symport-1 benzylhydantoin transporter from Microbacterium liquefaciens (Mhp1), we constructed a three-dimensional model of UreA which, combined with site-directed and classical random mutagenesis, led to the identification of amino acids important for UreA function. Our approach allowed us to suggest roles for these residues in the binding, recognition and translocation of urea, and in the sorting of UreA to the membrane. Residues W82, Y106, A110, T133, N275, D286, Y388, Y437 and S446, located in transmembrane helixes 2, 3, 7 and 11, were found to be involved in the binding, recognition and/or translocation of urea and the sorting of UreA to the membrane. Y106, A110, T133 and Y437 seem to play a role in substrate selectivity, while S446 is necessary for proper sorting of UreA to the membrane. Other amino acids identified by random classical mutagenesis (G99, R141, A163, G168 and P639) may be important for the basic transporter's structure, its proper folding or its correct traffic to the membrane.
Keywords: permease; sodium : solute symporter-family; structure–function relationships.
Figures


References
-
- McDonald MD, Smith CP, Walsh PJ. 2006. The physiology and evolution of urea transport in fishes. J. Membr. Biol. 212, 93–107. (doi:10.1007/s00232-006-0869-5) - DOI - PubMed
-
- Witte CP. 2011. Urea metabolism in plants. Plant Sci. 180, 431–438. (doi:10.1016/j.plantsci.2010.11.010) - DOI - PubMed
-
- Wagemaker MJ, Welboren W, van der Drift C, Jetten MS, Van Griensven LJ, Op den Camp HJ. 2005. The ornithine cycle enzyme arginase from Agaricus bisporus and its role in urea accumulation in fruit bodies. Biochim. Biophys. Acta 1681, 107–115. (doi:10.1016/j.bbaexp.2004.10.007) - DOI - PubMed
-
- Smith CP. 2009. Mammalian urea transporters. Exp. Physiol. 94, 180–185. (doi:10.1113/expphysiol.2008.043042) - DOI - PubMed
-
- Davis RH. 1970. Sources of urea in Neurospora. Biochim. Biophys. Acta 215, 412–414. (doi:10.1016/0304-4165(70)90042-5) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
