

Motor-mediated cortical versus astral microtubule organization in lipid-monolayered droplets
- PMID: 24966327
- PMCID: PMC4139258
- DOI: 10.1074/jbc.M114.582015
Motor-mediated cortical versus astral microtubule organization in lipid-monolayered droplets
Abstract
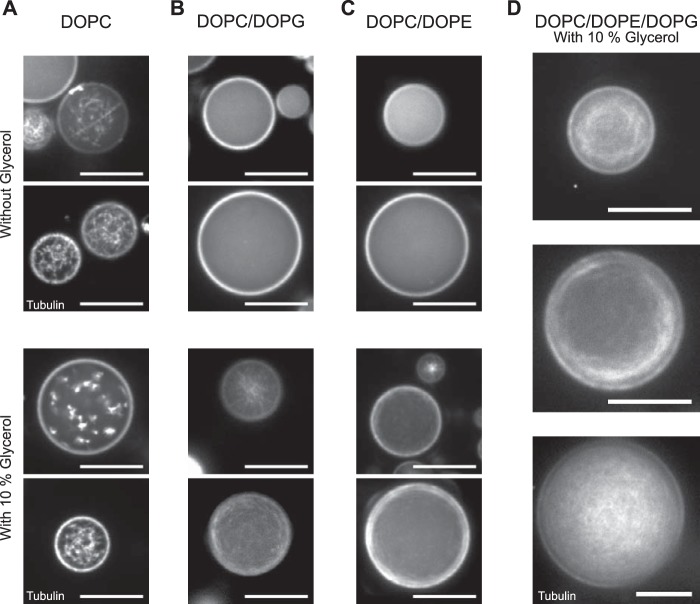
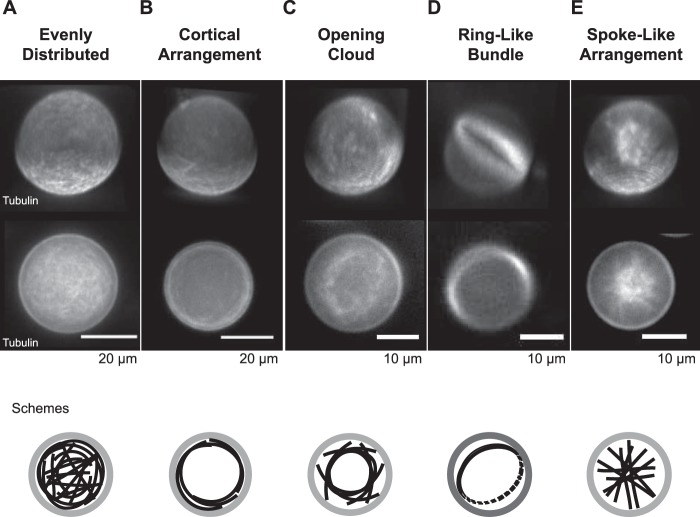
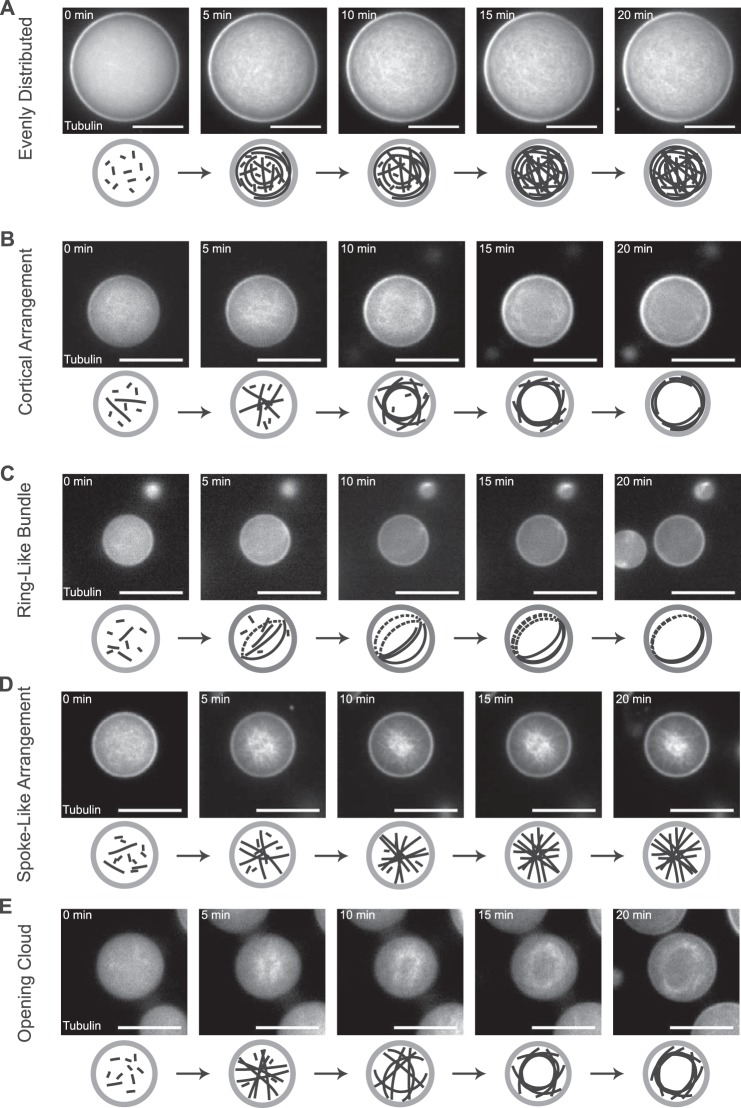
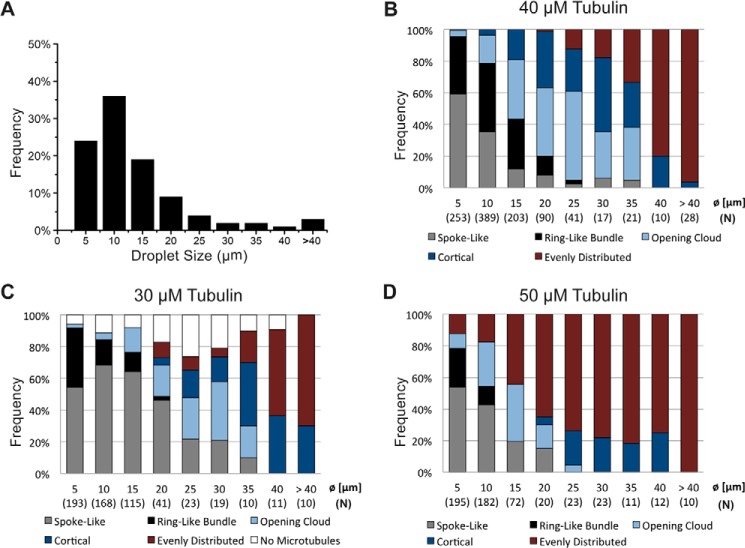
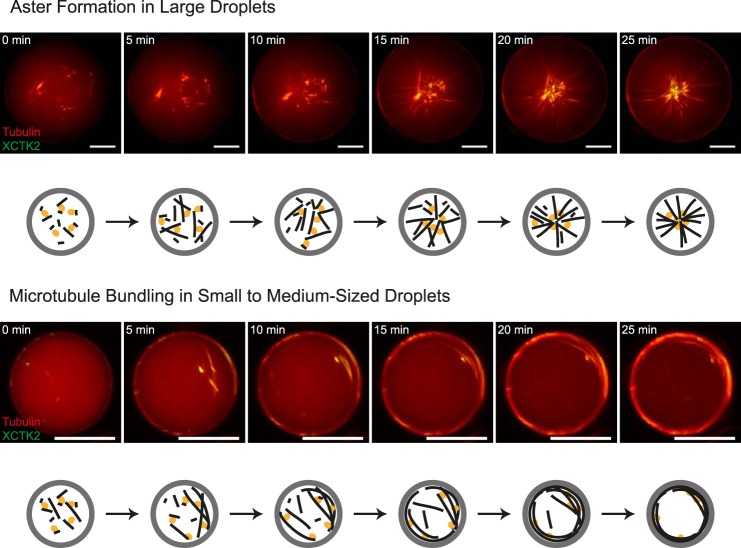
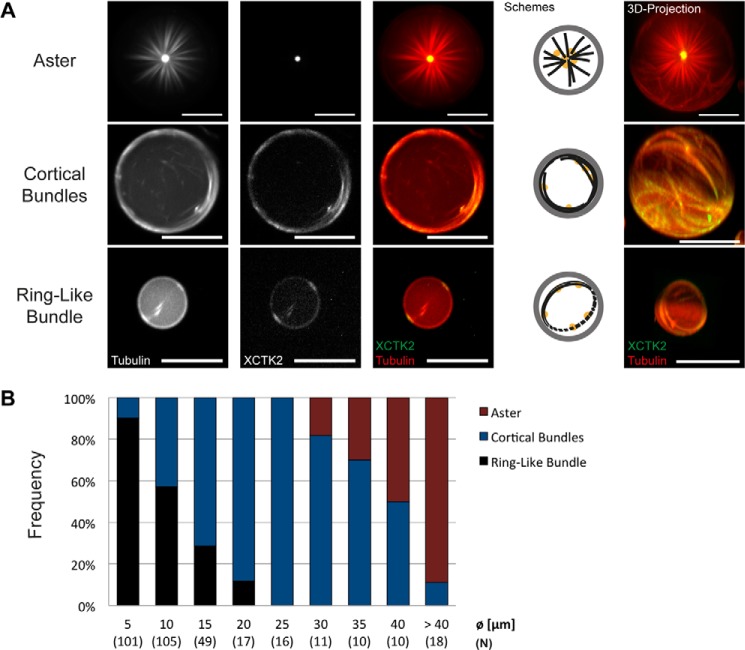
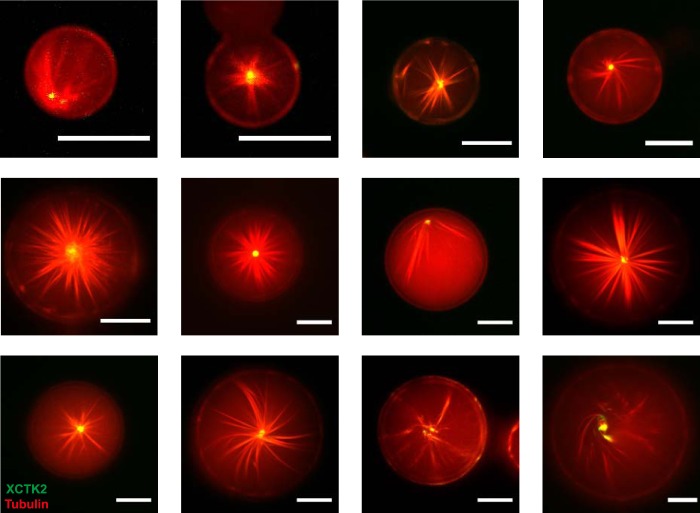
The correct spatial organization of microtubules is of crucial importance for determining the internal architecture of eukaryotic cells. Microtubules are arranged in space by a multitude of biochemical activities and by spatial constraints imposed by the cell boundary. The principles underlying the establishment of distinct intracellular architectures are only poorly understood. Here, we studied the effect of spatial confinement on the self-organization of purified motors and microtubules that are encapsulated in lipid-monolayered droplets in oil, varying in diameter from 5-100 μm, which covers the size range of typical cell bodies. We found that droplet size alone had a major organizing influence. The presence of a microtubule-crosslinking motor protein decreased the number of accessible types of microtubule organizations. Depending on the degree of spatial confinement, the presence of the motor caused either the formation of a cortical array of bent microtubule bundles or the generation of single microtubule asters in the droplets. These are two of the most prominent forms of microtubule arrangements in plant and metazoan cells. Our results provide insights into the combined organizing influence of spatial constraints and cross-linking motor activities determining distinct microtubule architectures in a minimal biomimetic system. In the future, this simple lipid-monolayered droplet system characterized here can be expanded readily to include further biochemical activities or used as the starting point for the investigation of motor-mediated microtubule organization inside liposomes surrounded by a deformable lipid bilayer.
Keywords: Confocal Microscopy; Cytoskeleton; Microtubule; Molecular Motor; Protein Self-Assembly; Reaction Compartmentalization; Self-organization.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Deinum E. E., Mulder B. M. (2013) Modelling the role of microtubules in plant cell morphology. Curr. Opin. Plant Biol. 16, 688–692 - PubMed
-
- Dogterom M., Surrey T. (2013) Microtubule organization in vitro. Curr. Opin. Cell Biol. 25, 23–29 - PubMed
-
- Helmke K. J., Heald R., Wilbur J. D. (2013) Interplay between spindle architecture and function. Int. Rev. Cell Mol. Biol. 306, 83–125 - PubMed
-
- Nédélec F. J., Surrey T., Maggs A. C., Leibler S. (1997) Self-organization of microtubules and motors. Nature 389, 305–308 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
