

Small ubiquitin-like modifier (SUMO) isoforms and conjugation-independent function in DNA double-strand break repair pathways
- PMID: 24966330
- PMCID: PMC4118090
- DOI: 10.1074/jbc.C114.582122
Small ubiquitin-like modifier (SUMO) isoforms and conjugation-independent function in DNA double-strand break repair pathways
Abstract
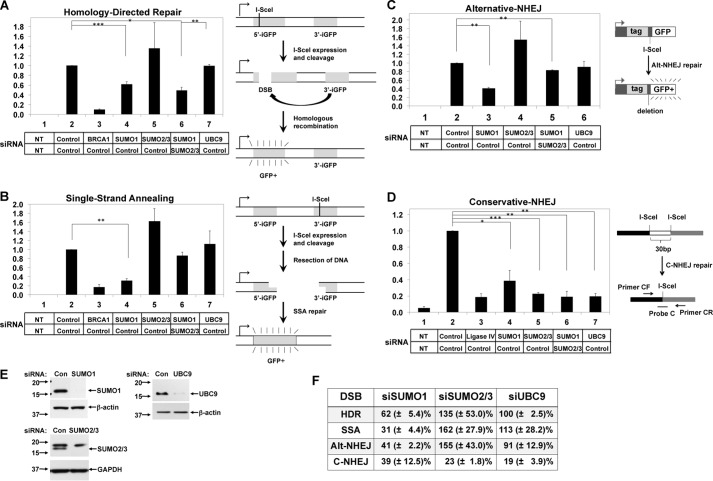
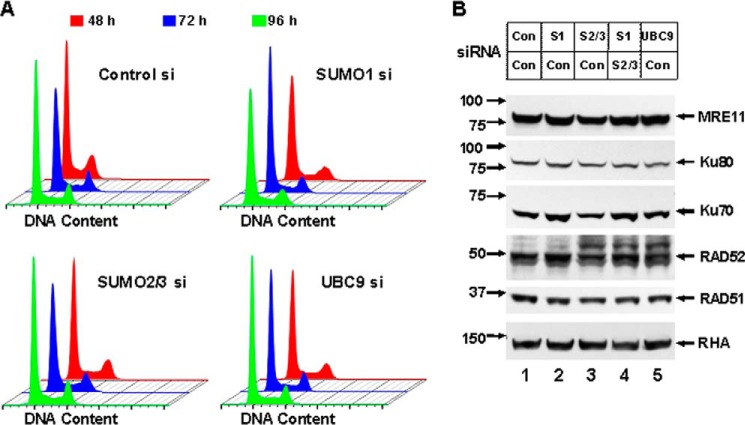
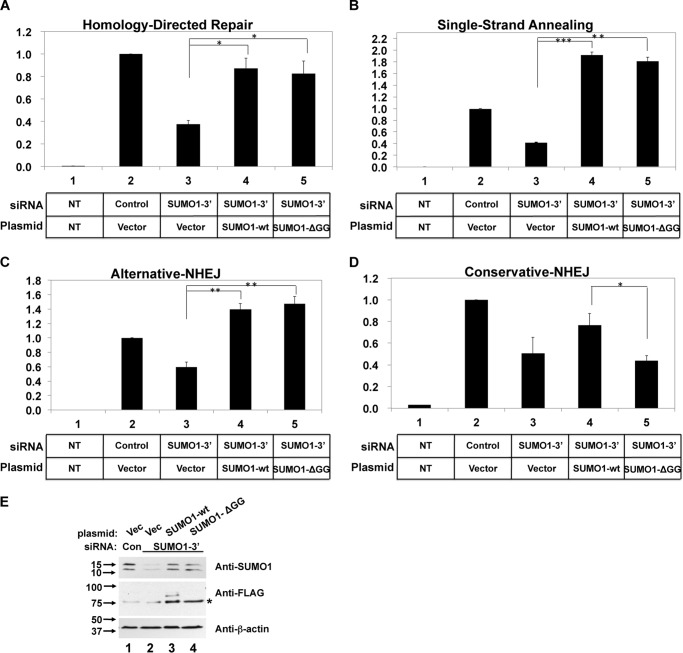
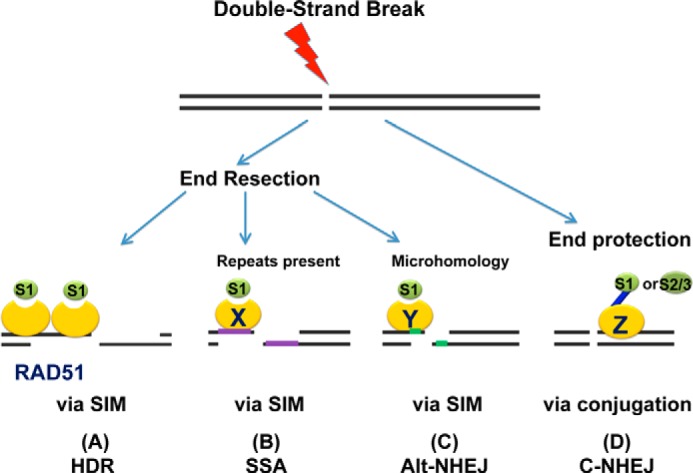
Small ubiquitin-like modifier (SUMO) proteins act in DNA double-strand break (DSB) repair, but the pathway specificity of the three major isoforms has not been defined. In experiments in which we depleted the endogenous SUMO protein by RNAi, we found that SUMO1 functioned in all subpathways of either homologous recombination (HR) or non-homologous end joining (NHEJ), whereas SUMO2/3 was required for the major NHEJ pathway, called conservative NHEJ, but dispensable in other DSB repair pathways. To our surprise, we found that depletion of UBC9, the unique SUMO E2 enzyme, had no effect in HR or alternative NHEJ (Alt-NHEJ) but was required for conservative NHEJ. Consistent with this result, both non-conjugatable mutant and wild-type SUMO1 proteins functioned similarly in HR and Alt-NHEJ. These results detail the functional roles of specific SUMO isoforms in DSB repair in mammalian cells and reveal that SUMO1 functions in HR or Alt-NHEJ as a free protein and not as a protein conjugate.
Keywords: DNA Damage Response; Double-strand Break Repair; Homologous Recombination; Non-homologous End Joining; SUMO-interacting Motif (SIM); Small Ubiquitin-like Modifier (SUMO); Sumoylation; UBC9.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Bunting S. F., Callén E., Wong N., Chen H. T., Polato F., Gunn A., Bothmer A., Feldhahn N., Fernandez-Capetillo O., Cao L., Xu X., Deng C. X., Finkel T., Nussenzweig M., Stark J. M., Nussenzweig A. (2010) 53BP1 inhibits homologous recombination in Brca1-deficient cells by blocking resection of DNA breaks. Cell 141, 243–254 - PMC - PubMed
-
- Daley J. M., Palmbos P. L., Wu D., Wilson T. E. (2005) Nonhomologous end joining in yeast. Annu. Rev. Genet. 39, 431–451 - PubMed
-
- Burma S., Chen B. P., Chen D. J. (2006) Role of non-homologous end joining (NHEJ) in maintaining genomic integrity. DNA Repair 5, 1042–1048 - PubMed
-
- Guirouilh-Barbat J., Huck S., Bertrand P., Pirzio L., Desmaze C., Sabatier L., Lopez B. S. (2004) Impact of the KU80 pathway on NHEJ-induced genome rearrangements in mammalian cells. Mol. Cell 14, 611–623 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
