

Porcine reproductive and respiratory syndrome virus induces IL-1β production depending on TLR4/MyD88 pathway and NLRP3 inflammasome in primary porcine alveolar macrophages
- PMID: 24966466
- PMCID: PMC4055429
- DOI: 10.1155/2014/403515
Porcine reproductive and respiratory syndrome virus induces IL-1β production depending on TLR4/MyD88 pathway and NLRP3 inflammasome in primary porcine alveolar macrophages
Abstract
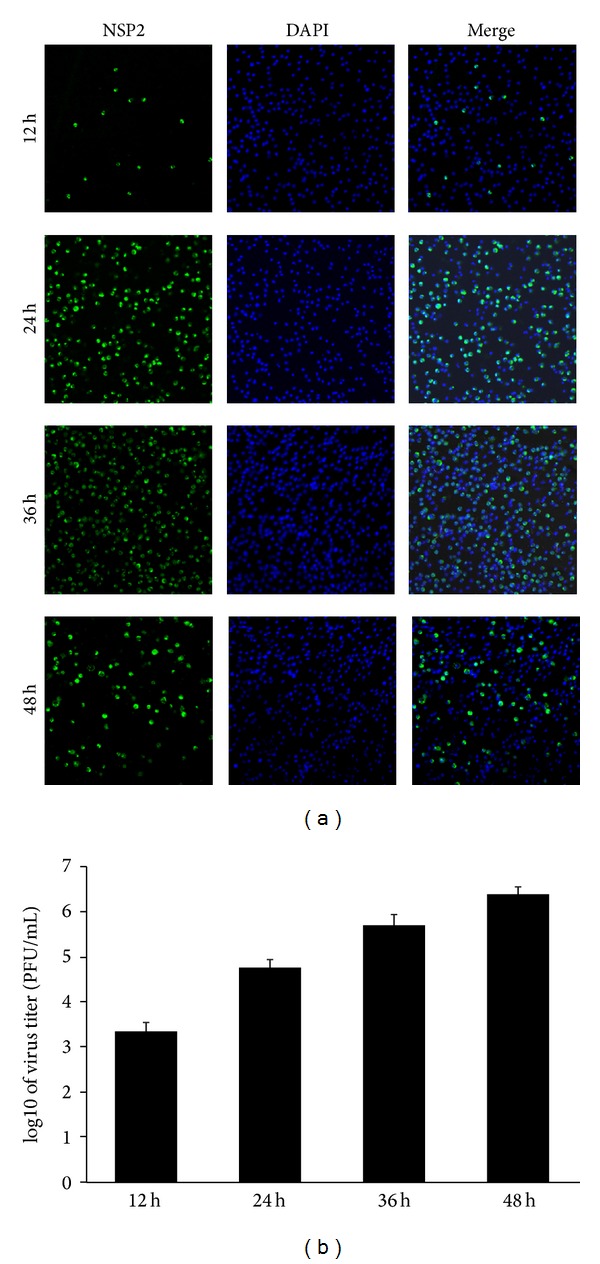
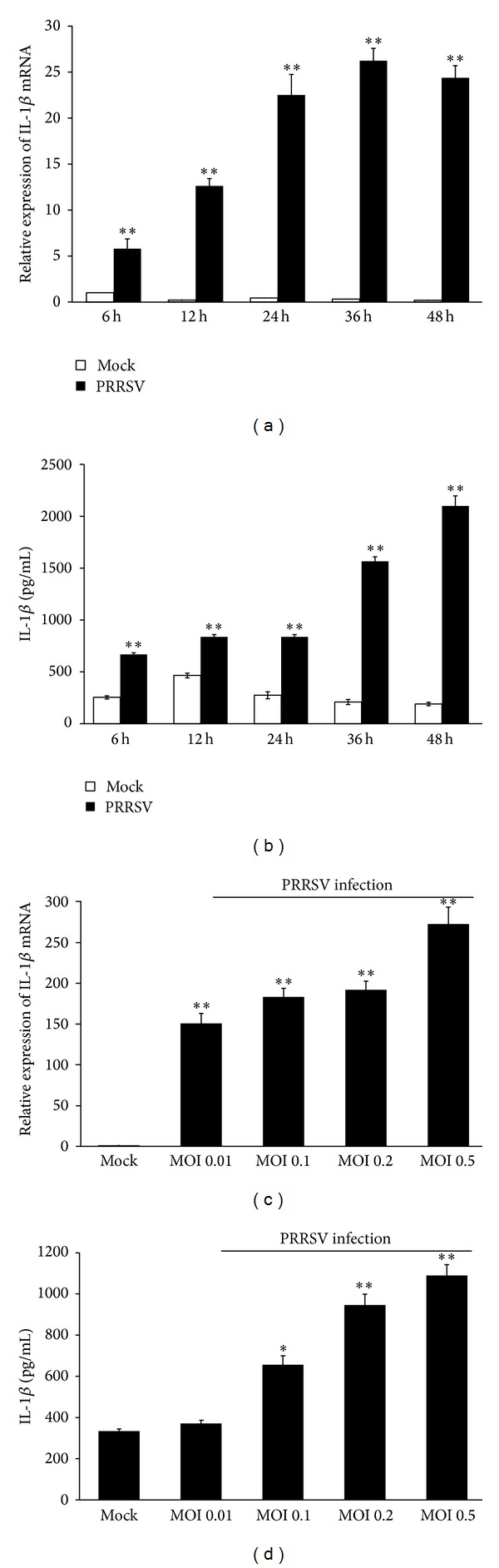
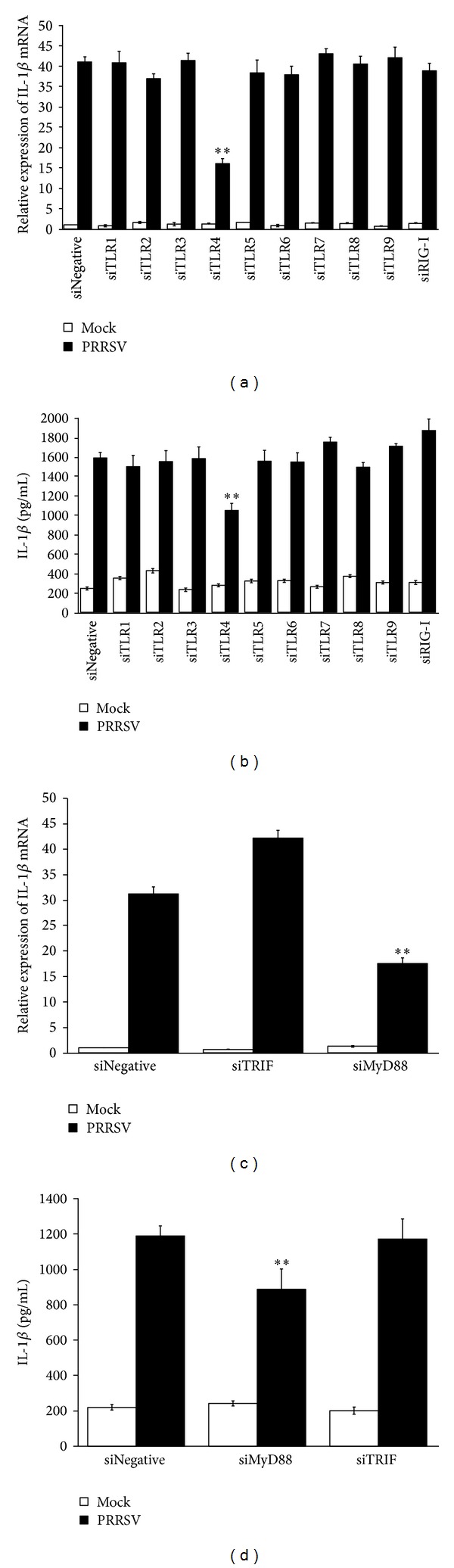
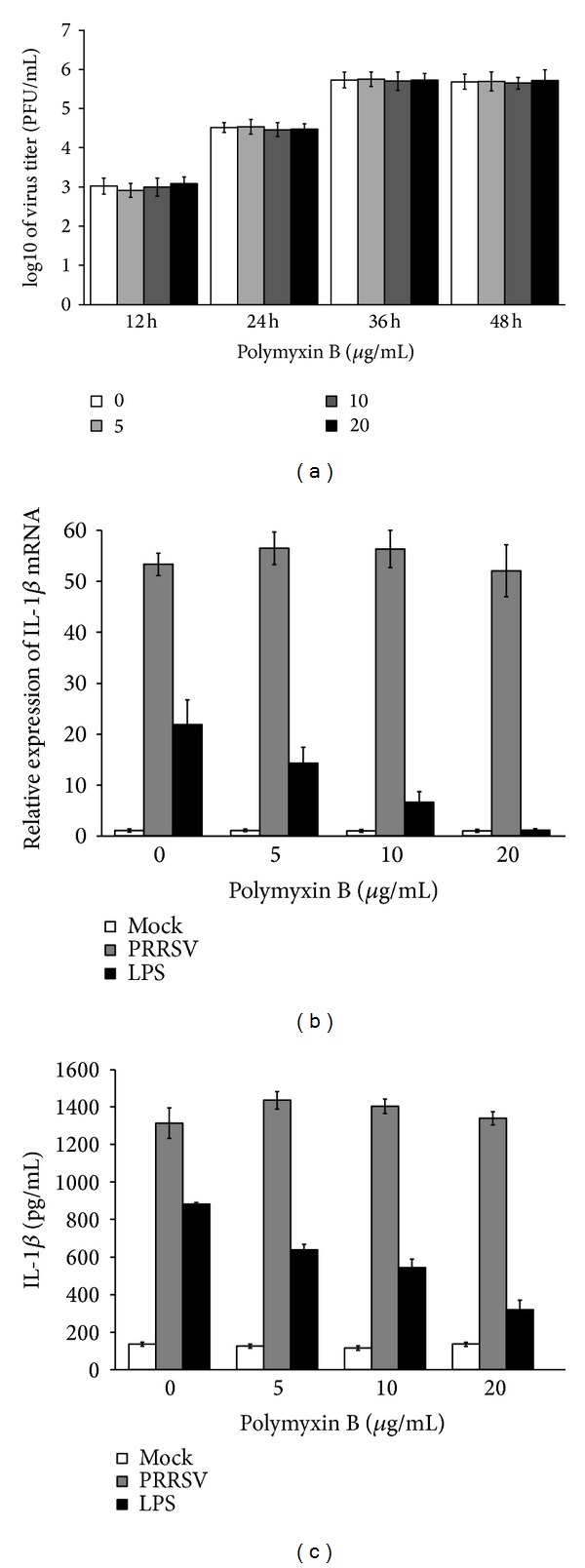
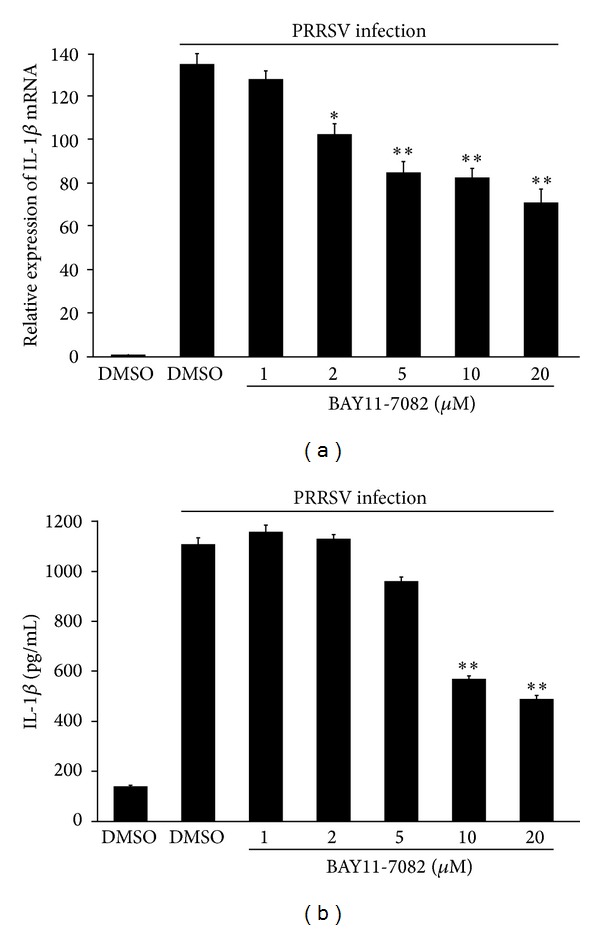
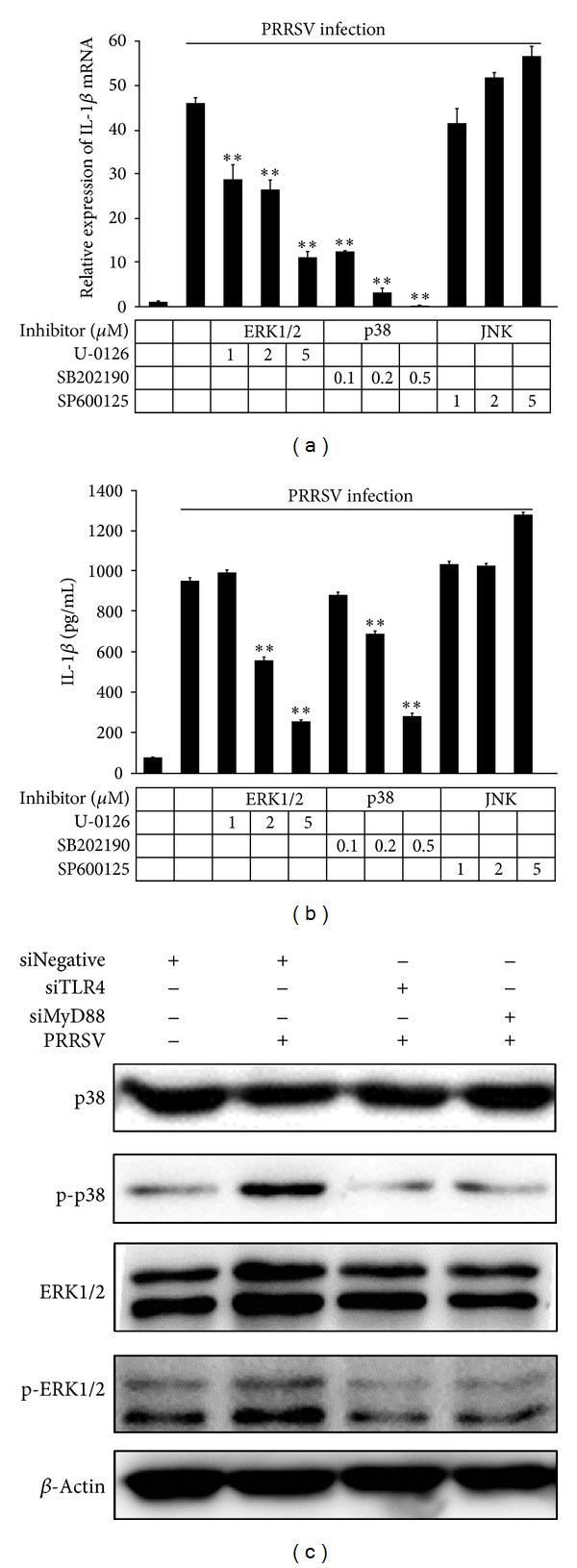
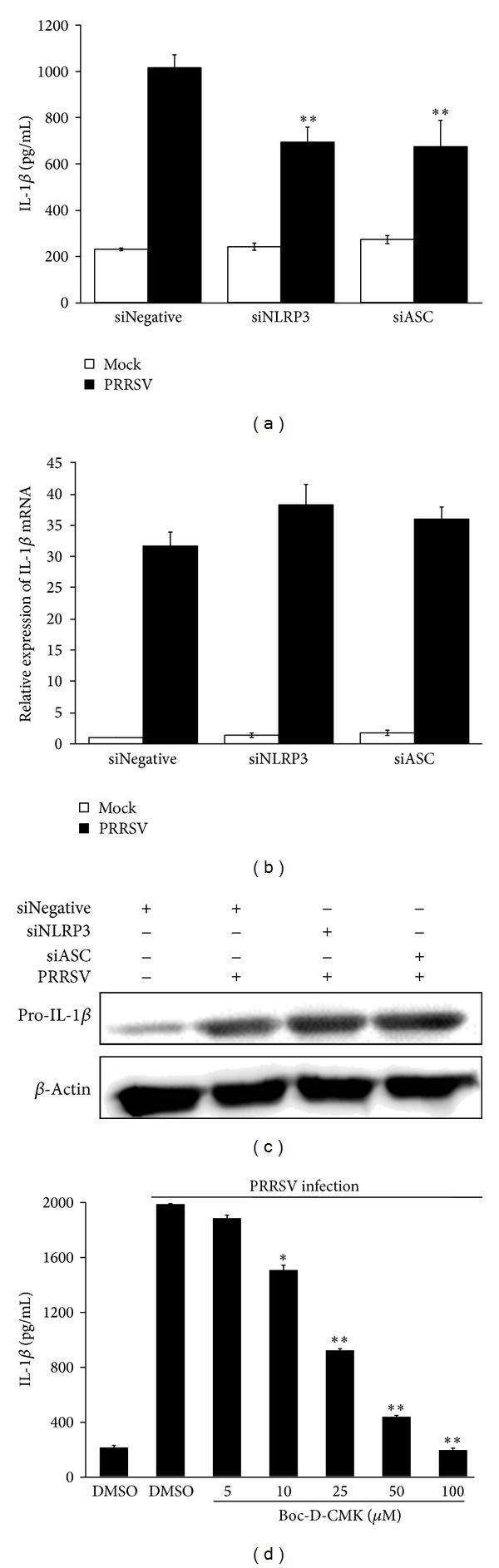
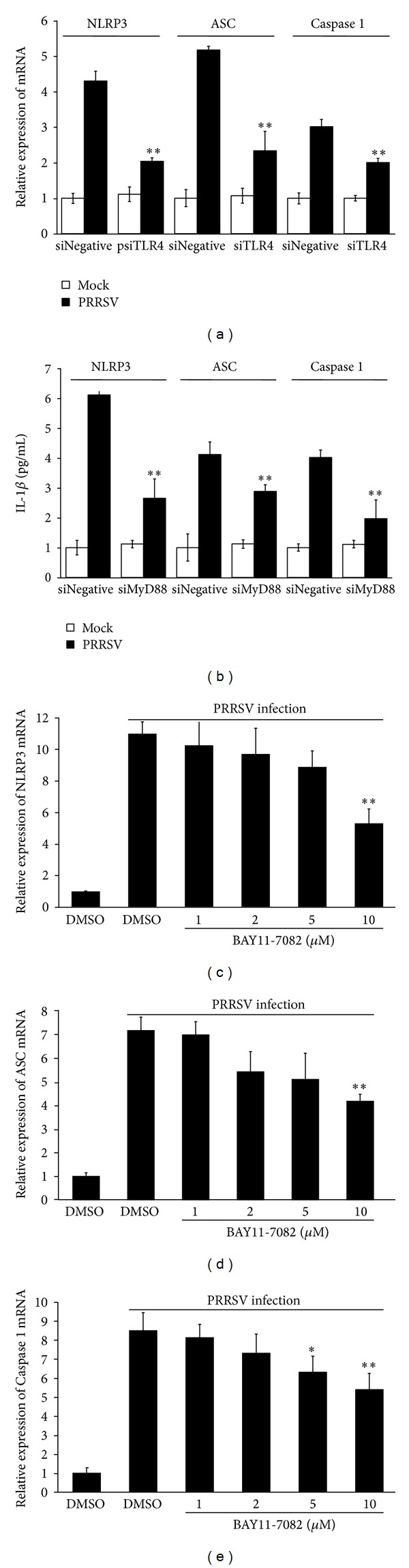
Porcine reproductive and respiratory syndrome virus (PRRSV) is an Arterivirus that has been devastating the swine industry worldwide since the late 1980s. Previous studies have reported that PRRSV infection induced the production of IL-1 β . However, the cellular sensors and signaling pathways involved in this process have not been elucidated yet. Here, we studied the mechanisms responsible for the production of IL-1 β in response to highly pathogenic PRRSV. Upon PRRSV infection of primary porcine alveolar macrophages, both mRNA expression and secretion of IL-1 β were significantly increased in a time- and dose-dependent manner. We also investigated the role of several pattern-recognition receptors and adaptor molecules in this response and showed that the TLR4/MyD88 pathway and its downstream signaling molecules, NF- κ B, ERK1/2, and p38 MAPKs, were involved in IL-1 β production during PRRSV infection. Treatment with specific inhibitors or siRNA knockdown assays demonstrated that components of the NLRP3 inflammasome were crucial for IL-1 β secretion but not for IL-1 β mRNA expression. Furthermore, TLR4/MyD88/NF- κ B signaling pathway was involved in PRRSV-induced expression of NLRP3 inflammasome components. Together, our results deciphered the pathways leading from recognition of PRRSV to the production and release of IL-1 β , providing a deeper knowledge of the mechanisms of PRRSV-induced inflammation responses.
Figures
References
-
- Bose S, Banerjee AK. Innate immune response against nonsegmented negative strand RNA viruses. Journal of Interferon and Cytokine Research. 2003;23(8):401–412. - PubMed
-
- Dinarello CA. Biologic basis for Interleukin-1 in disease. Blood. 1996;87(6):2095–2147. - PubMed
-
- Eder C. Mechanisms of interleukin-1β release. Immunobiology. 2009;214(7):543–553. - PubMed
-
- Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
