

Converging on a core cognitive deficit: the impact of various neurodevelopmental insults on cognitive control
- PMID: 24966811
- PMCID: PMC4052340
- DOI: 10.3389/fnins.2014.00153
Converging on a core cognitive deficit: the impact of various neurodevelopmental insults on cognitive control
Abstract
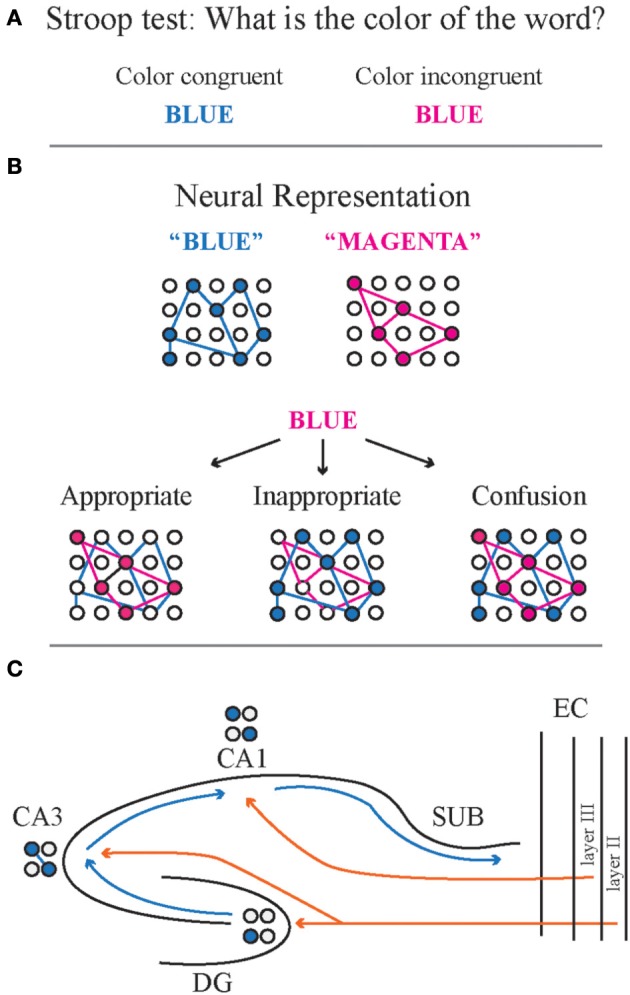
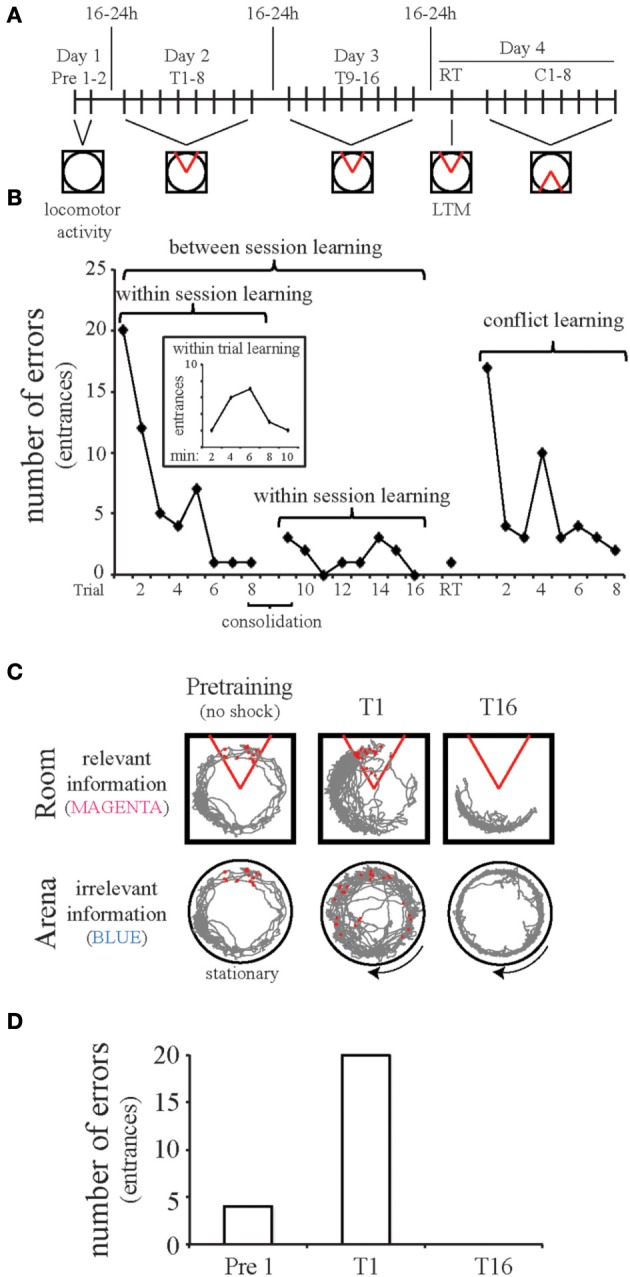
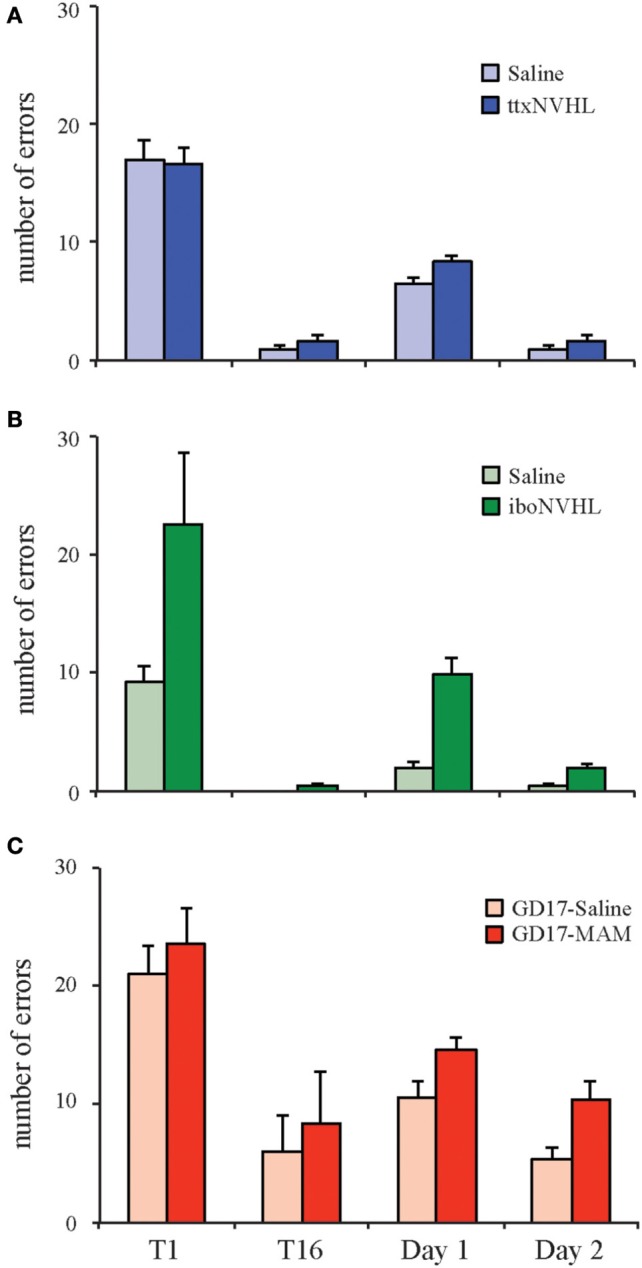
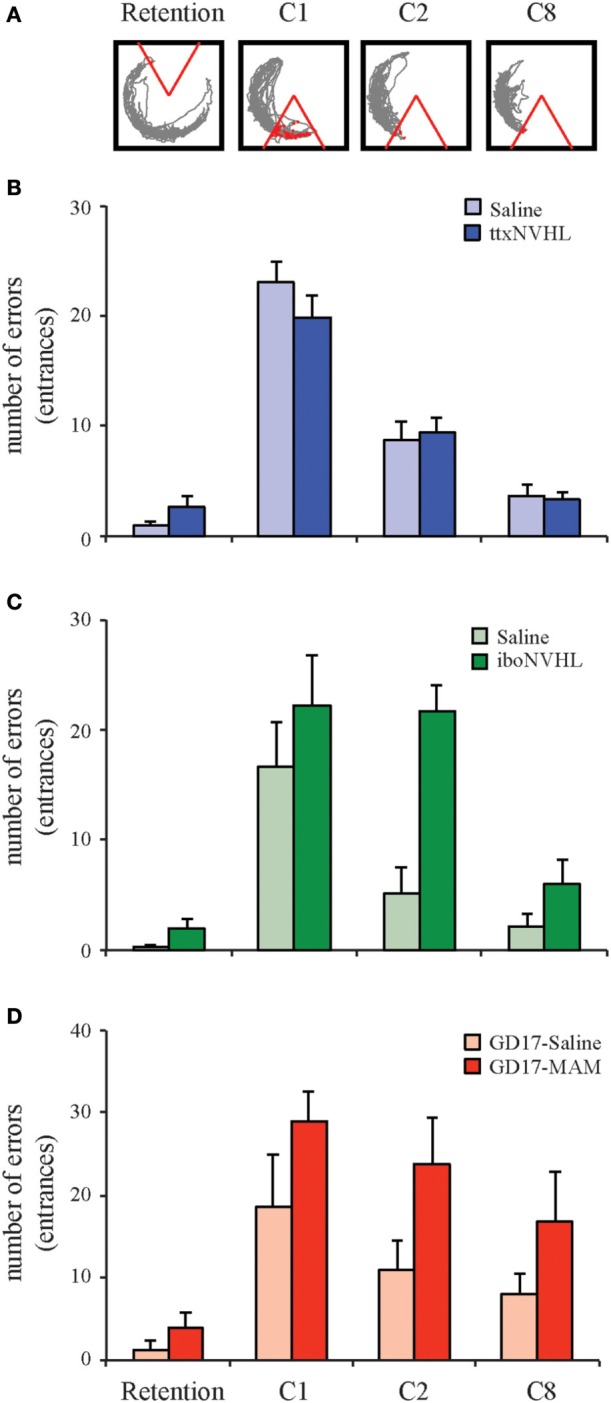
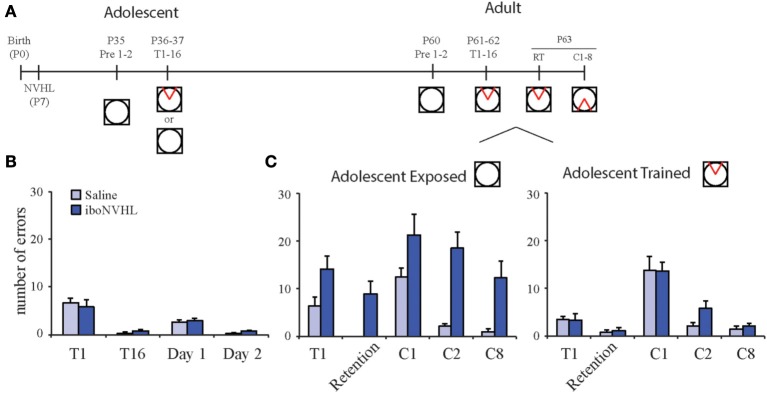
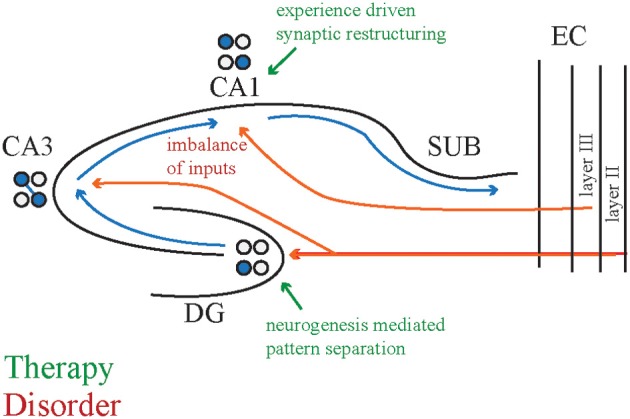
Despite substantial effort and immense need, the treatment options for major neuropsychiatric illnesses like schizophrenia are limited and largely ineffective at improving the most debilitating cognitive symptoms that are central to mental illness. These symptoms include cognitive control deficits, the inability to selectively use information that is currently relevant and ignore what is currently irrelevant. Contemporary attempts to accelerate progress are in part founded on an effort to reconceptualize neuropsychiatric illness as a disorder of neural development. This neuro-developmental framework emphasizes abnormal neural circuits on the one hand, and on the other, it suggests there are therapeutic opportunities to exploit the developmental processes of excitatory neuron pruning, inhibitory neuron proliferation, elaboration of myelination, and other circuit refinements that extend through adolescence and into early adulthood. We have crafted a preclinical research program aimed at cognition failures that may be relevant to mental illness. By working with a variety of neurodevelopmental rodent models, we strive to identify a common pathophysiology that underlies cognitive control failure as well as a common strategy for improving cognition in the face of neural circuit abnormalities. Here we review our work to characterize cognitive control deficits in rats with a neonatal ventral hippocampus lesion and rats that were exposed to Methylazoxymethanol acetate (MAM) in utero. We review our findings as they pertain to early developmental processes, including neurogenesis, as well as the power of cognitive experience to refine neural circuit function within the mature and maturing brain's cognitive circuitry.
Keywords: cognitive control; hippocampus; mental illness; neural coordination; neurodevelopmental insults; schizophrenia models.
Figures
References
-
- Bachevalier J., Beauregard M. (1993). Maturation of medial temporal lobe memory functions in rodents, monkeys, and humans. Hippocampus 3, 191–201 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources