

Vimentin mediates uptake of C3 exoenzyme
- PMID: 24967582
- PMCID: PMC4072758
- DOI: 10.1371/journal.pone.0101071
Vimentin mediates uptake of C3 exoenzyme
Abstract
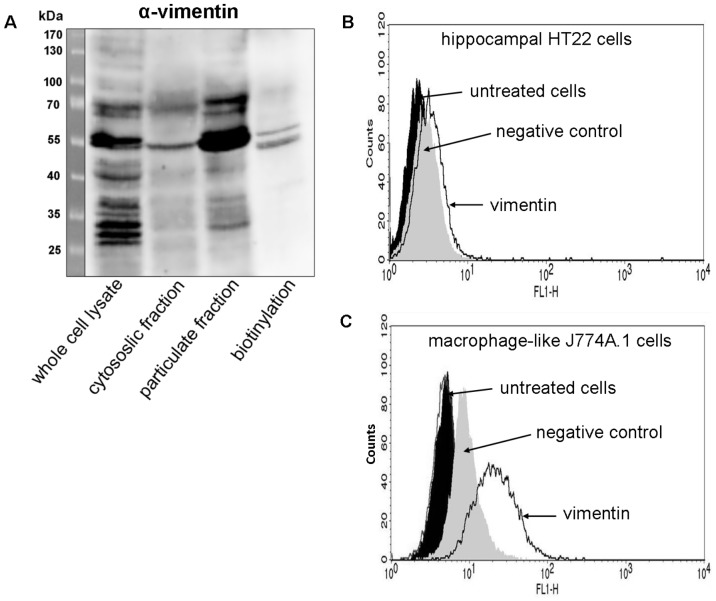
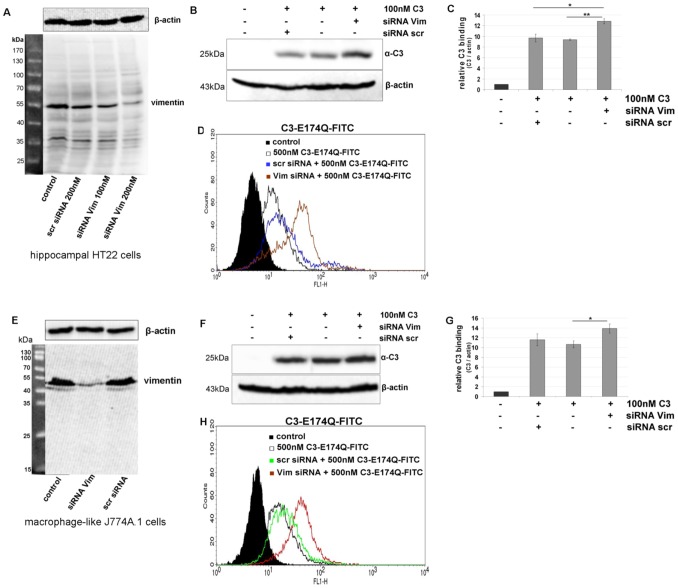
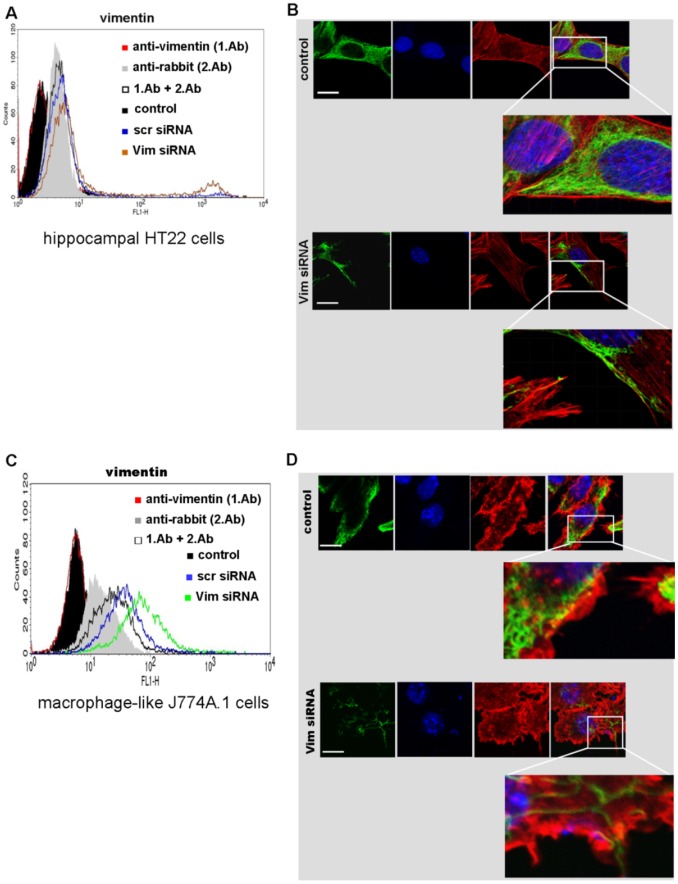
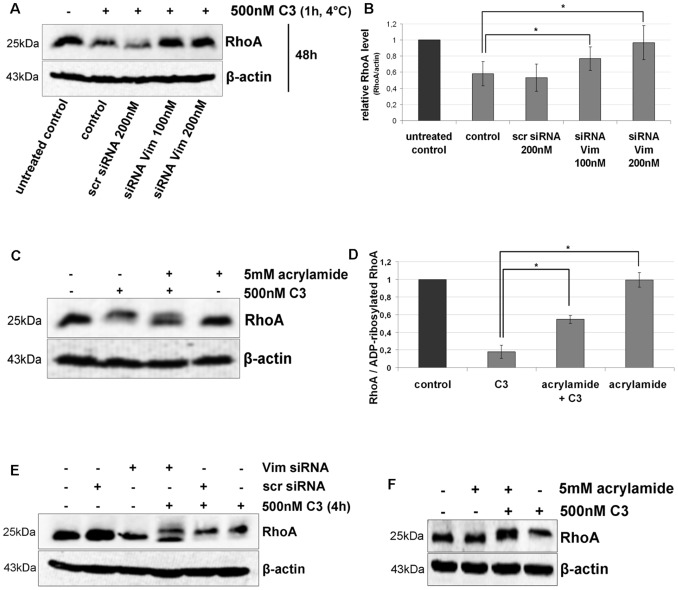
Clostridium botulinum C3 exoenzyme (C3) selectively inactivates RhoA/B/C GTPases by ADP-ribosylation. Based on this substrate specificity C3 is a well-established tool in cell biology. C3 is taken up by eukaryotic cells although lacking an uptake and translocation domain. Based on different approaches vimentin was identified as membranous C3-interaction partner by mass spectrometry. Vimentin in fact was partly localized at the outer surface of hippocampal HT22 cells and J744A.1 macrophages. Domain analysis identified the rod domain as binding partner of C3. Vimentin was also involved in uptake of C3 as shown by knock down of vimentin in HT22 and J774A.1 cells. The involvement of vimentin in uptake of C3 was further supported by the findings that the vimentin disruptor acrylamide blocked uptake of C3. Vimentin is not only a major organizing element of the intermediate filament network but is also involved in both binding and uptake of C3 exoenzyme.
Conflict of interest statement
Figures
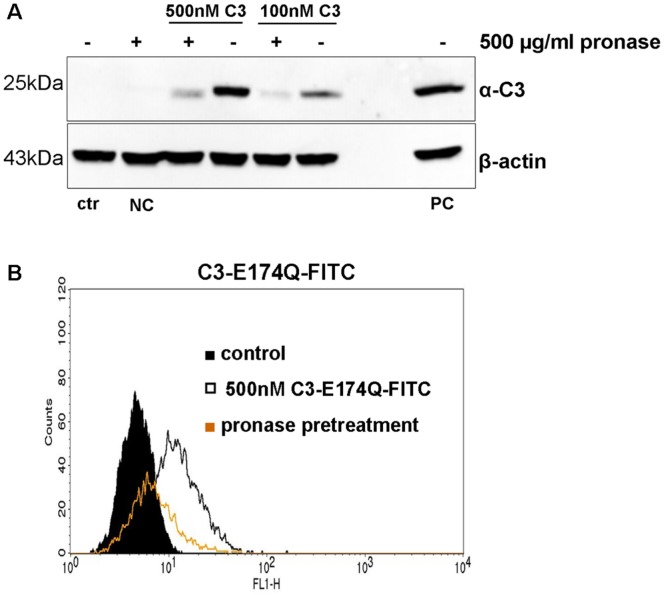
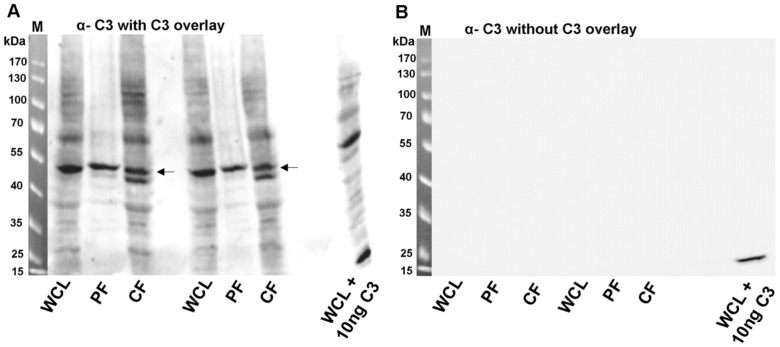
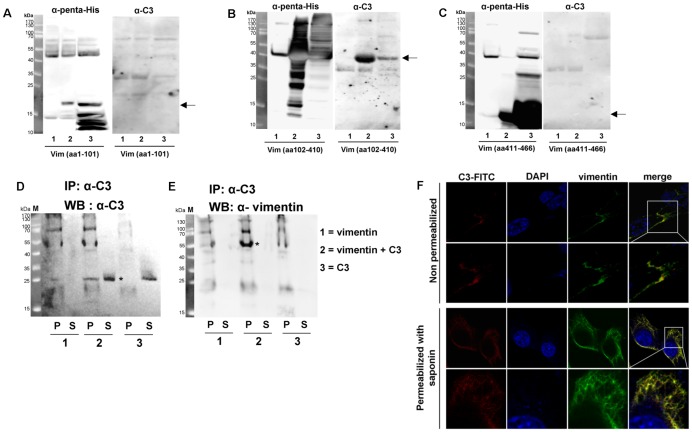
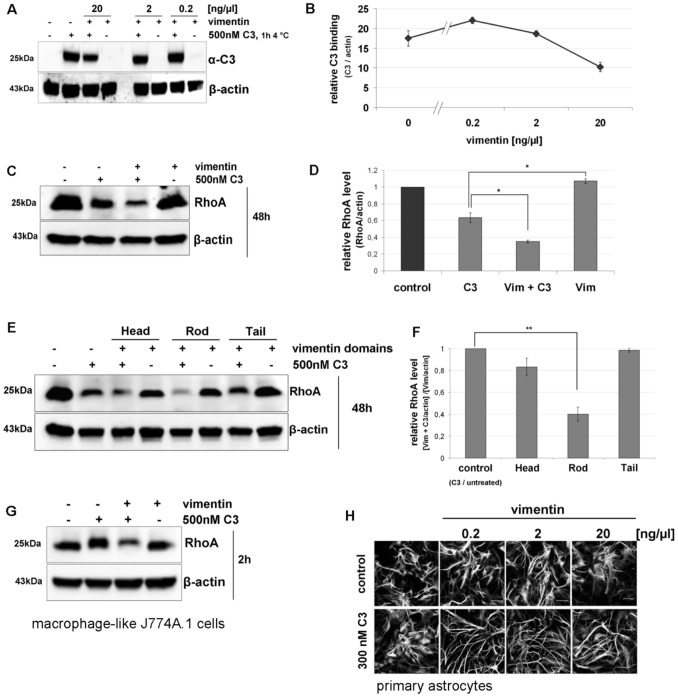
Similar articles
-
Uptake of clostridium botulinum C3 exoenzyme into intact HT22 and J774A.1 cells.Toxins (Basel). 2015 Feb 2;7(2):380-95. doi: 10.3390/toxins7020380. Toxins (Basel). 2015. PMID: 25648844 Free PMC article.
-
Cell Entry of C3 Exoenzyme from Clostridium botulinum.Curr Top Microbiol Immunol. 2017;406:97-118. doi: 10.1007/82_2016_44. Curr Top Microbiol Immunol. 2017. PMID: 27832378 Free PMC article. Review.
-
Binding of Clostridium botulinum C3 exoenzyme to intact cells.Naunyn Schmiedebergs Arch Pharmacol. 2014 Jun;387(6):523-32. doi: 10.1007/s00210-014-0963-7. Epub 2014 Mar 2. Naunyn Schmiedebergs Arch Pharmacol. 2014. PMID: 24584821
-
The intermediate filament protein vimentin is essential for axonotrophic effects of Clostridium botulinum C3 exoenzyme.J Neurochem. 2016 Oct;139(2):234-244. doi: 10.1111/jnc.13739. Epub 2016 Aug 9. J Neurochem. 2016. PMID: 27419376
-
Conformational plasticity is crucial for C3-RhoA complex formation by ARTT-loop.Pathog Dis. 2015 Dec;73(9):ftv094. doi: 10.1093/femspd/ftv094. Epub 2015 Oct 15. Pathog Dis. 2015. PMID: 26474844 Review.
Cited by
-
Secreted Phospholipases A2 - not just Enzymes: Revisited.Int J Biol Sci. 2022 Jan 1;18(2):873-888. doi: 10.7150/ijbs.68093. eCollection 2022. Int J Biol Sci. 2022. PMID: 35002531 Free PMC article. Review.
-
Some Examples of Bacterial Toxins as Tools.Toxins (Basel). 2024 Apr 23;16(5):202. doi: 10.3390/toxins16050202. Toxins (Basel). 2024. PMID: 38787054 Free PMC article. Review.
-
Vimentin as a Multifaceted Player and Potential Therapeutic Target in Viral Infections.Int J Mol Sci. 2020 Jun 30;21(13):4675. doi: 10.3390/ijms21134675. Int J Mol Sci. 2020. PMID: 32630064 Free PMC article. Review.
-
Lentiviral Vector-Mediated Expression of Exoenzyme C3 Transferase Lowers Intraocular Pressure in Monkeys.Mol Ther. 2019 Jul 3;27(7):1327-1338. doi: 10.1016/j.ymthe.2019.04.021. Epub 2019 May 9. Mol Ther. 2019. PMID: 31129118 Free PMC article.
-
A SARS-CoV-2 neutralizing antibody protects from lung pathology in a COVID-19 hamster model.bioRxiv [Preprint]. 2020 Aug 16:2020.08.15.252320. doi: 10.1101/2020.08.15.252320. bioRxiv. 2020. Update in: Cell. 2020 Nov 12;183(4):1058-1069.e19. doi: 10.1016/j.cell.2020.09.049. PMID: 32817952 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
