

Ly6C(high) monocytes become alternatively activated macrophages in schistosome granulomas with help from CD4+ cells
- PMID: 24967715
- PMCID: PMC4072804
- DOI: 10.1371/journal.ppat.1004080
Ly6C(high) monocytes become alternatively activated macrophages in schistosome granulomas with help from CD4+ cells
Abstract
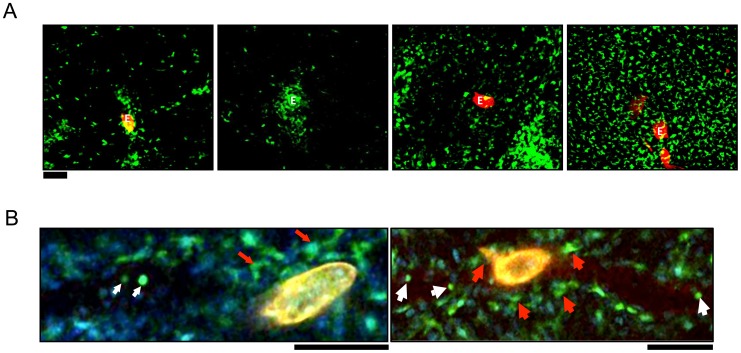
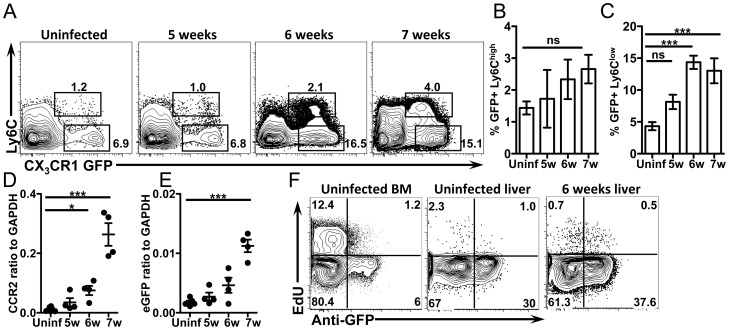
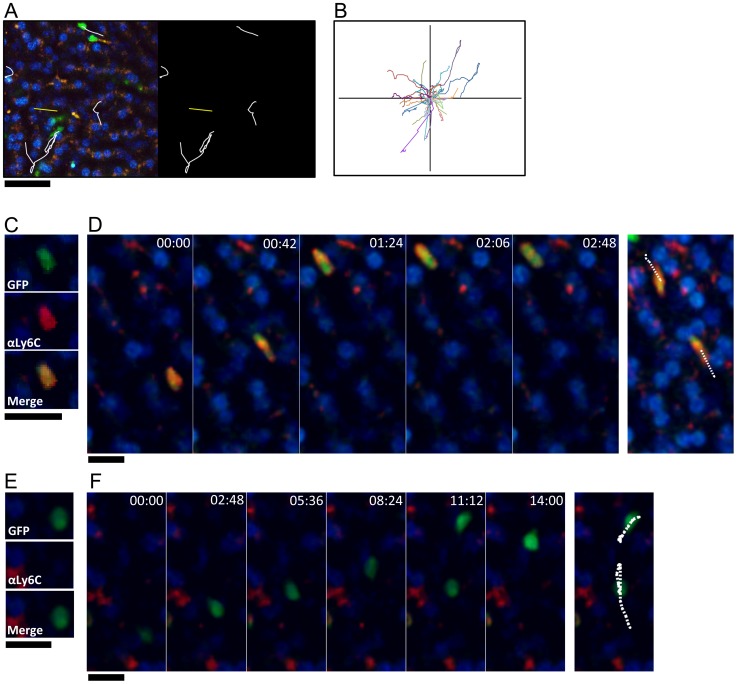
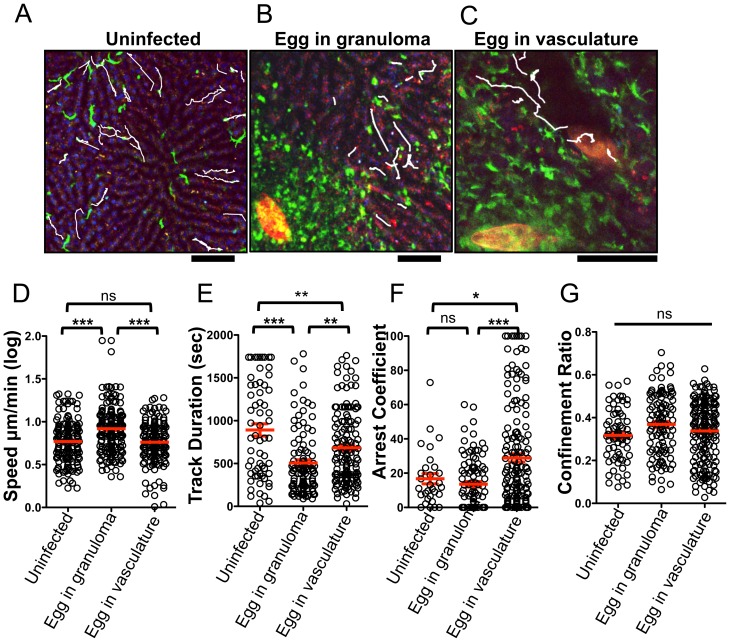
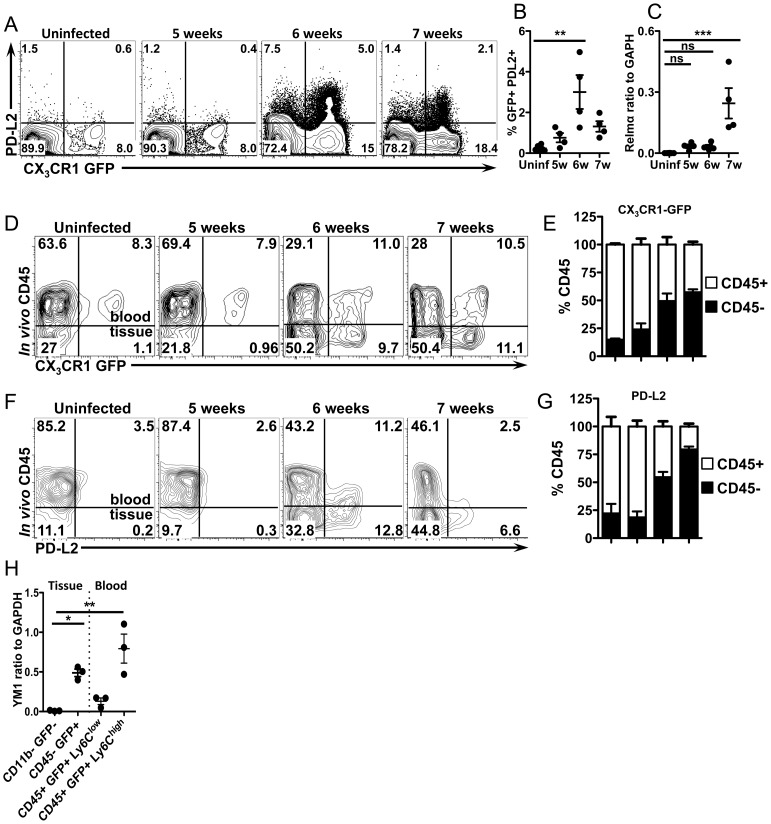
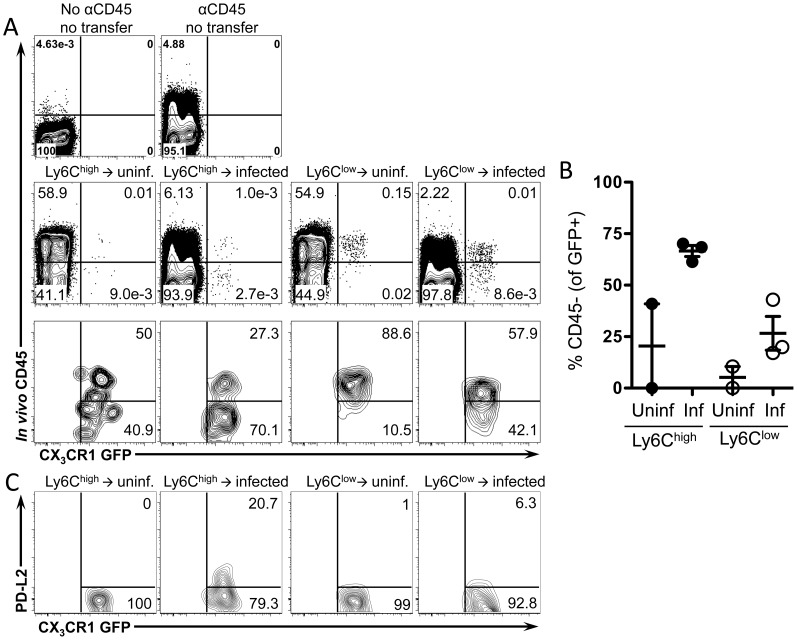
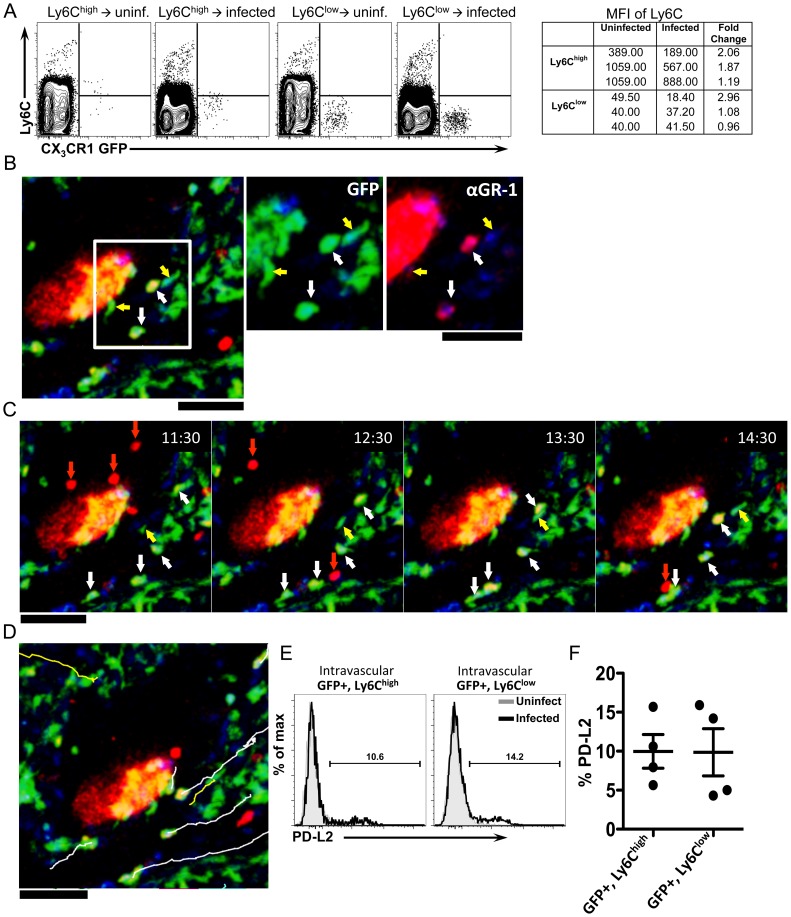
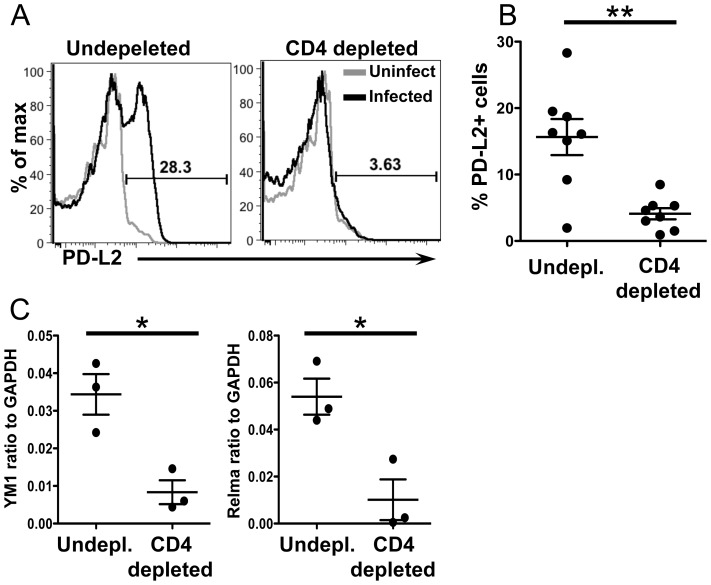
Alternatively activated macrophages (AAM) that accumulate during chronic T helper 2 inflammatory conditions may arise through proliferation of resident macrophages or recruitment of monocyte-derived cells. Liver granulomas that form around eggs of the helminth parasite Schistosoma mansoni require AAM to limit tissue damage. Here, we characterized monocyte and macrophage dynamics in the livers of infected CX3CR1(GFP/+) mice. CX₃CR1-GFP⁺ monocytes and macrophages accumulated around eggs and in granulomas during infection and upregulated PD-L2 expression, indicating differentiation into AAM. Intravital imaging of CX₃CR1-GFP⁺ Ly6C(low) monocytes revealed alterations in patrolling behavior including arrest around eggs that were not encased in granulomas. Differential labeling of CX₃CR1-GFP⁺ cells in the blood and the tissue showed CD4⁺ T cell dependent accumulation of PD-L2⁺ CX₃CR1-GFP⁺ AAM in the tissues as granulomas form. By adoptive transfer of Ly6C(high) and Ly6C(low) monocytes into infected mice, we found that AAM originate primarily from transferred Ly6C(high) monocytes, but that these cells may transition through a Ly6C(low) state and adopt patrolling behavior in the vasculature. Thus, during chronic helminth infection AAM can arise from recruited Ly6C(high) monocytes via help from CD4⁺ T cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Allen JE, Maizels RM (2011) Diversity and dialogue in immunity to helminths. Nat Rev Immunol 11: 375–388. - PubMed
-
- Gordon S, Martinez FO (2010) Alternative activation of macrophages: mechanism and functions. Immunity 32: 593–604. - PubMed
-
- Herbert DR, Holscher C, Mohrs M, Arendse B, Schwegmann A, et al. (2004) Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity 20: 623–635. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
