

A nucleic-acid hydrolyzing single chain antibody confers resistance to DNA virus infection in hela cells and C57BL/6 mice
- PMID: 24968358
- PMCID: PMC4072776
- DOI: 10.1371/journal.ppat.1004208
A nucleic-acid hydrolyzing single chain antibody confers resistance to DNA virus infection in hela cells and C57BL/6 mice
Erratum in
- PLoS Pathog. 2014 Jul;10(7):e1004318
Abstract
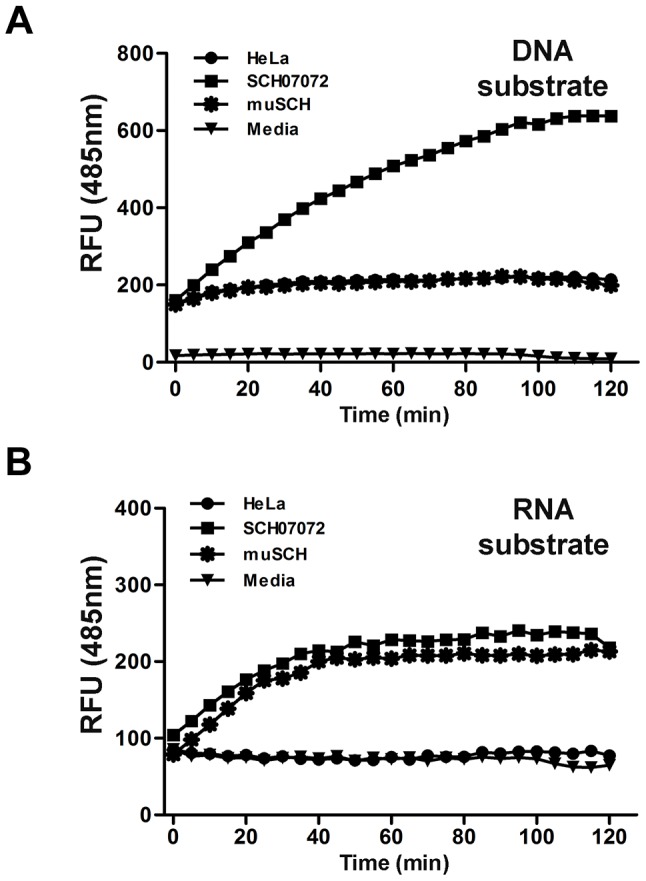
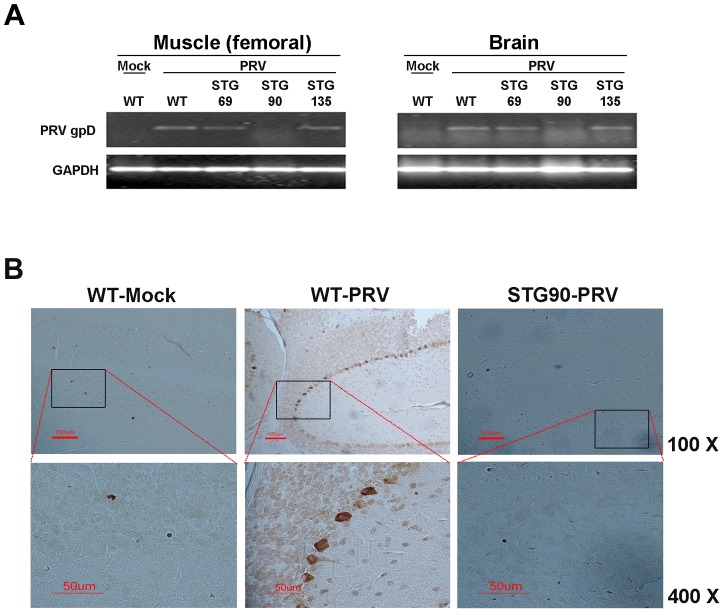
Viral protein neutralizing antibodies have been developed but they are limited only to the targeted virus and are often susceptible to antigenic drift. Here, we present an alternative strategy for creating virus-resistant cells and animals by ectopic expression of a nucleic acid hydrolyzing catalytic 3D8 single chain variable fragment (scFv), which has both DNase and RNase activities. HeLa cells (SCH07072) [corrected] expressing 3D8 scFv acquired significant resistance to DNA viruses. Virus challenging with Herpes simplex virus (HSV) in 3D8 scFv transgenic cells and fluorescence resonance energy transfer (FRET) assay based on direct DNA cleavage analysis revealed that the induced resistance in HeLa cells was acquired by the nucleic acid hydrolyzing catalytic activity of 3D8 scFv. In addition, pseudorabies virus (PRV) infection in WT C57BL/6 mice was lethal, whereas transgenic mice (STG90) that expressed high levels of 3D8 scFv mRNA in liver, muscle, and brain showed a 56% survival rate 5 days after PRV intramuscular infection. The antiviral effects against DNA viruses conferred by 3D8 scFv expression in HeLa cells as well as an in vivo mouse system can be attributed to the nuclease activity that inhibits viral genome DNA replication in the nucleus and/or viral mRNA translation in the cytoplasm. Our results demonstrate that the nucleic-acid hydrolyzing activity of 3D8 scFv confers viral resistance to DNA viruses in vitro in HeLa cells and in an in vivo mouse system.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
-
- Ding S (2008) Virology: Principles and Applications. The Yale Journal of Biology and Medicine 81: 155.
-
- Osterhaus A (2009) Timely tale of avian flu. Nature 462: 986.
-
- Yamada T, Dautry A, Walport M (2008) Ready for avian flu? Nature 454: 162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
