

Dissection of the adenoviral VA RNAI central domain structure reveals minimum requirements for RNA-mediated inhibition of PKR
- PMID: 24970889
- PMCID: PMC4132820
- DOI: 10.1074/jbc.M114.550046
Dissection of the adenoviral VA RNAI central domain structure reveals minimum requirements for RNA-mediated inhibition of PKR
Abstract
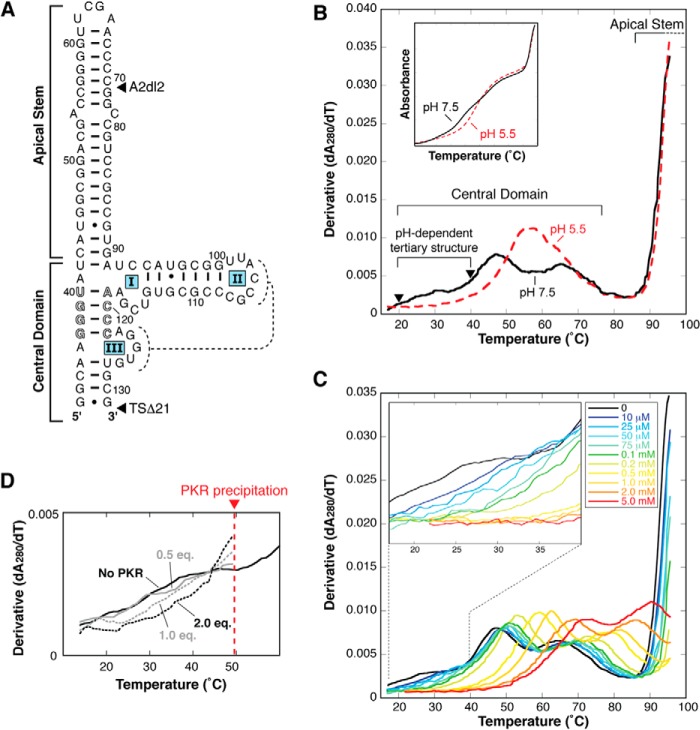
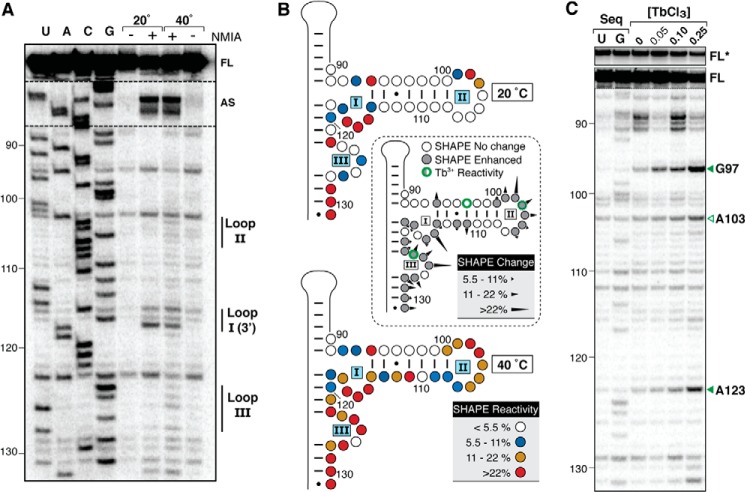
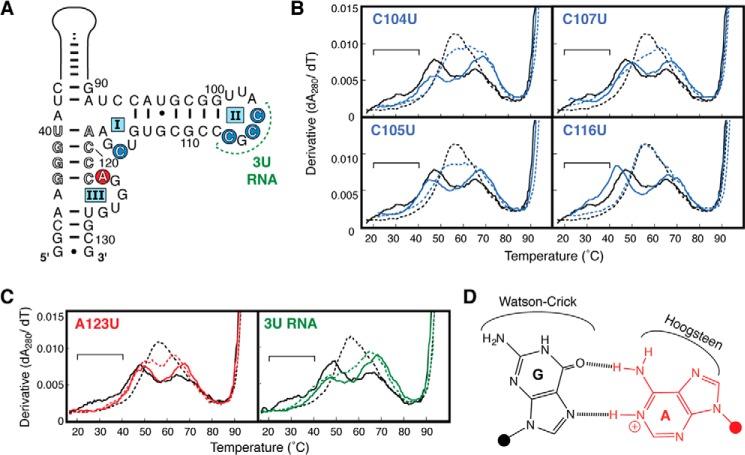
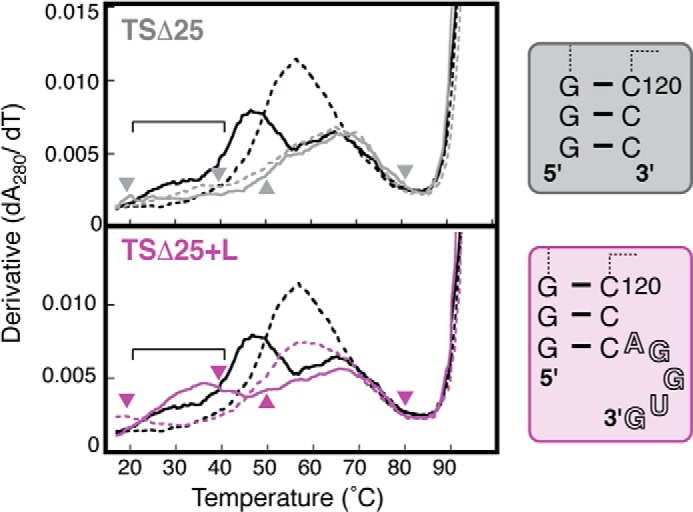
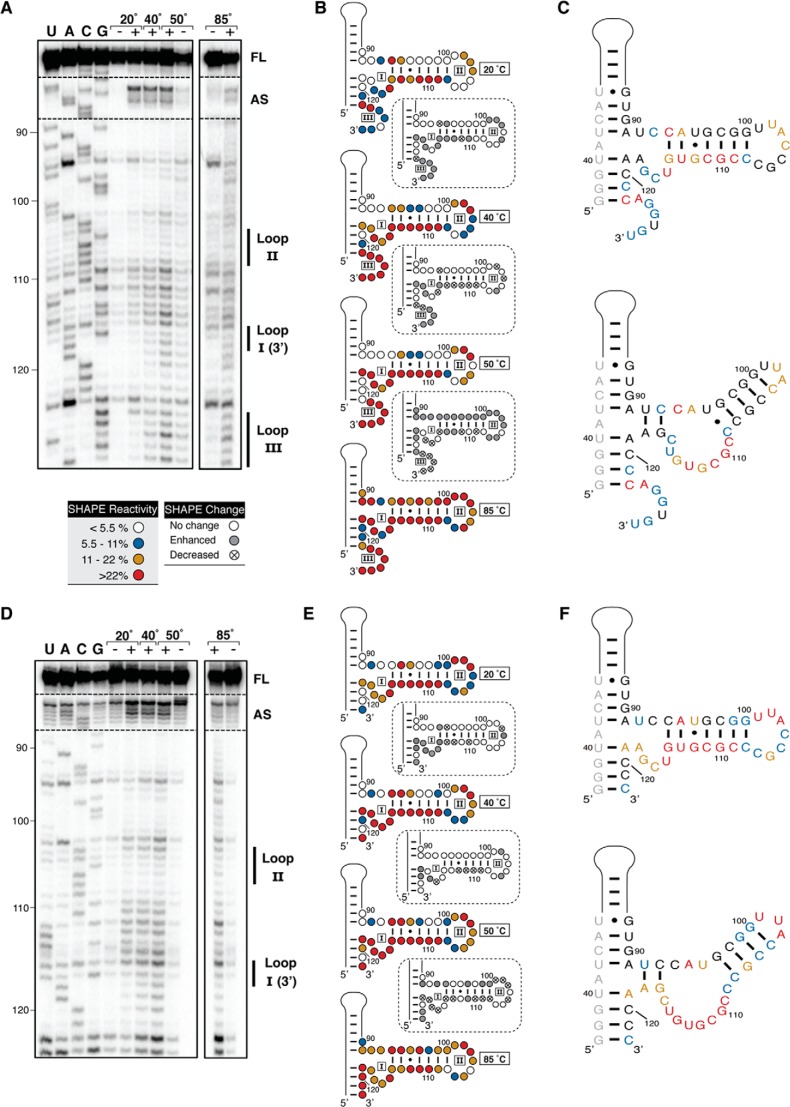
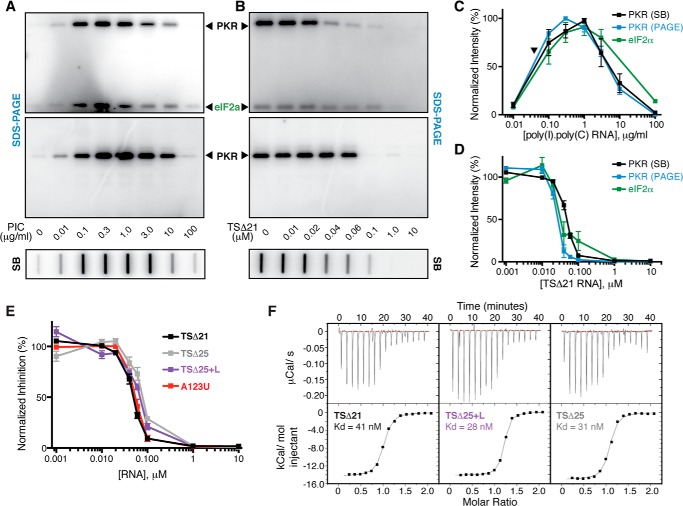
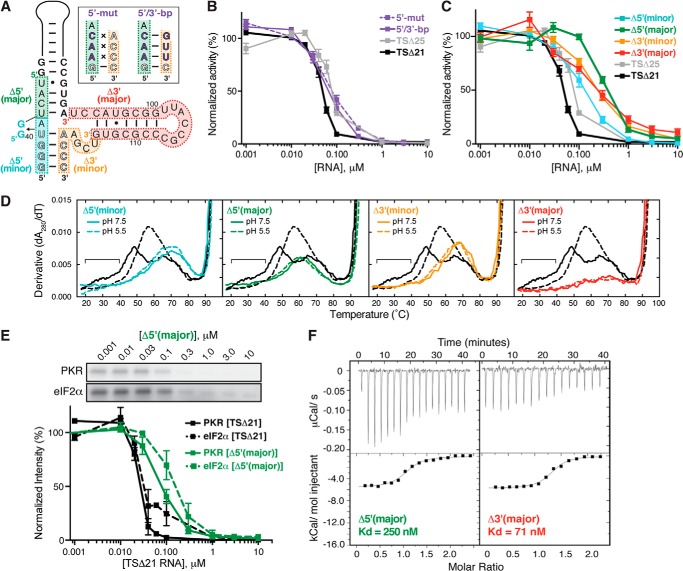
Virus-associated RNA I (VA RNAI) is a short (∼160-nucleotide) non-coding RNA transcript employed by adenoviruses to subvert the innate immune system protein double-stranded RNA-activated protein kinase (PKR). The central domain of VA RNAI is proposed to contain a complex tertiary structure that contributes to its optimal inhibitory activity against PKR. Here we use a combination of VA RNAI mutagenesis, structural analyses, as well as PKR activity and binding assays to dissect this tertiary structure and assess its functional role. Our results support the existence of a pH- and Mg(2+)-dependent tertiary structure involving pseudoknot formation within the central domain. Unexpectedly, this structure appears to play no direct role in PKR inhibition. Deletion of central domain sequences within a minimal but fully active construct lacking the tertiary structure reveals a crucial role in PKR binding and inhibition for nucleotides in the 5' half of the central domain. Deletion of the central domain 3' half also significantly impacts activity but appears to arise indirectly by reducing its capacity to assist in optimally presenting the 5' half sequence. Collectively, our results identify regions of VA RNAI critical for PKR inhibition and reveal that the requirements for an effective RNA inhibitor of PKR are simpler than appreciated previously.
Keywords: Adenovirus; Isothermal Titration Calorimetry (ITC); Mutagenesis; Protein Kinase RNA-activated (PKR); RNA Structure; RNA-Protein Interaction; SHAPE Structure Probing; Translation Control; UV Melting.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
