

4'-Ethynyl-2-fluoro-2'-deoxyadenosine (EFdA) inhibits HIV-1 reverse transcriptase with multiple mechanisms
- PMID: 24970894
- PMCID: PMC4148878
- DOI: 10.1074/jbc.M114.562694
4'-Ethynyl-2-fluoro-2'-deoxyadenosine (EFdA) inhibits HIV-1 reverse transcriptase with multiple mechanisms
Abstract
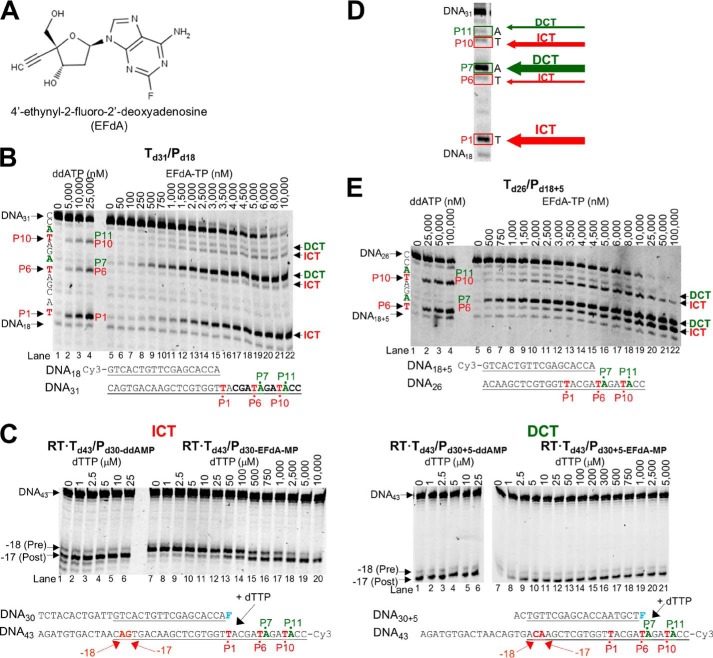
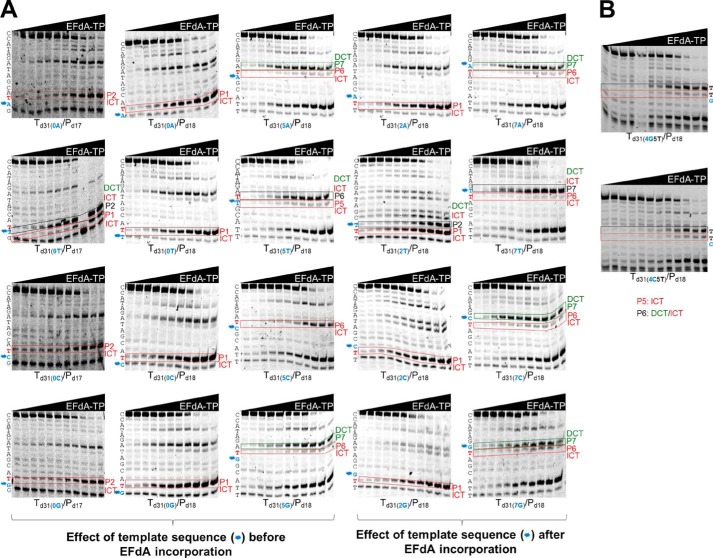
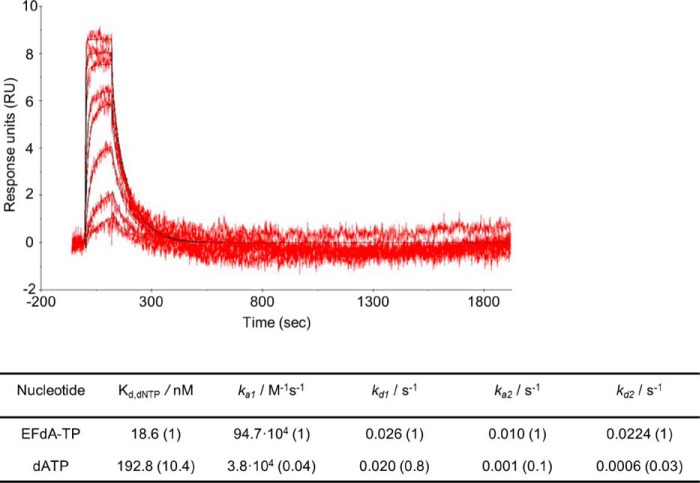
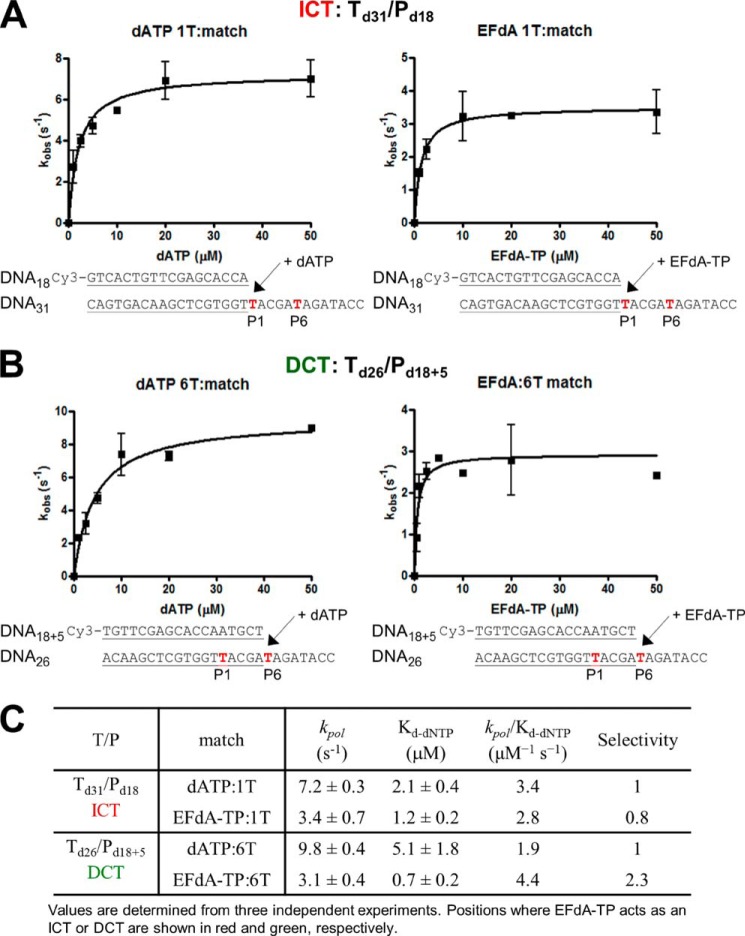
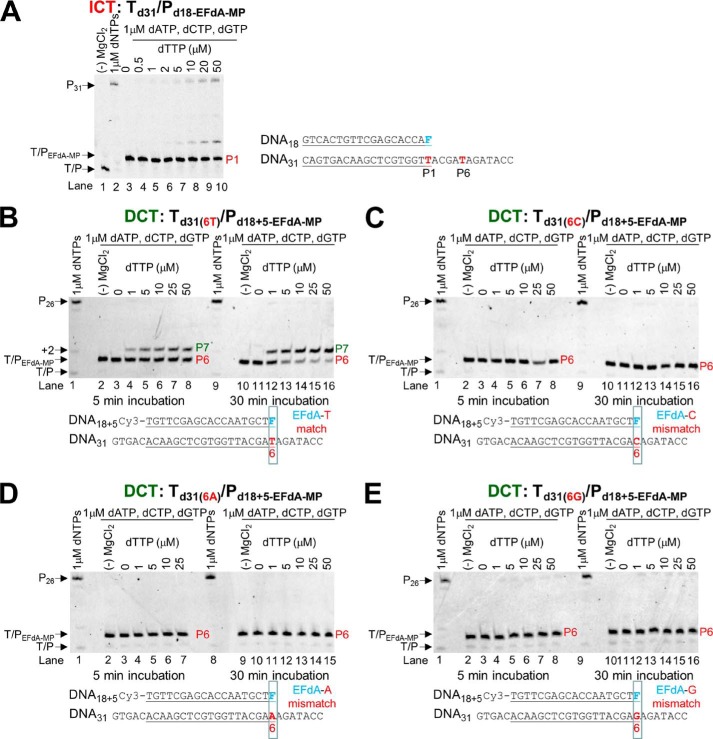
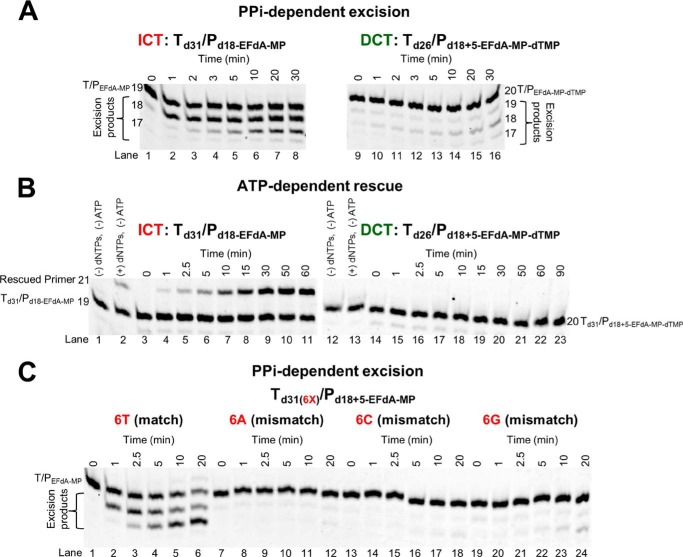
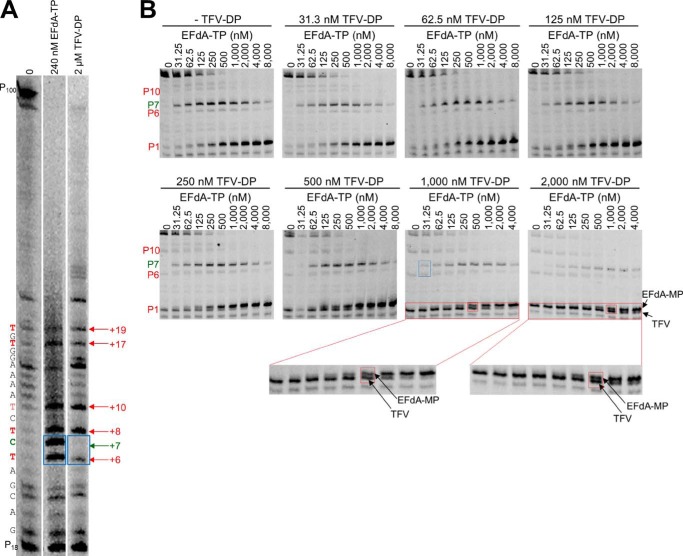
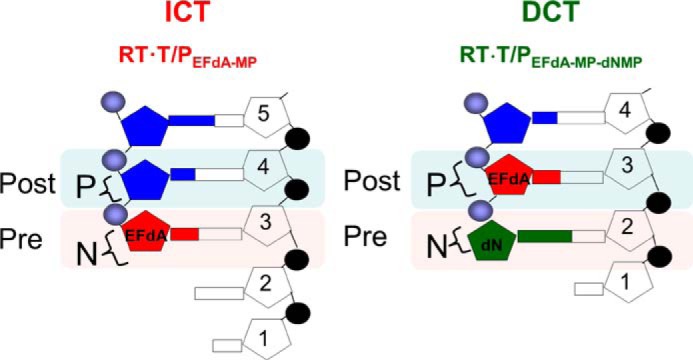
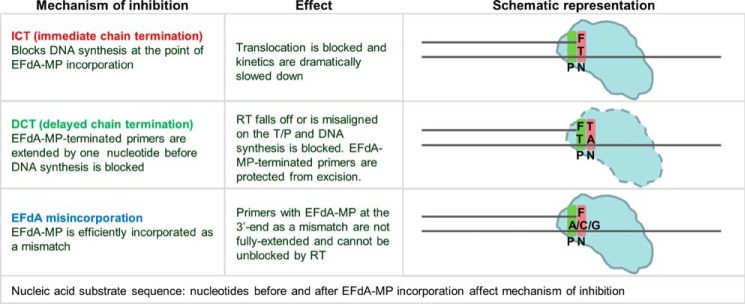
4'-Ethynyl-2-fluoro-2'-deoxyadenosine (EFdA) is a nucleoside analog that, unlike approved anti-human immunodeficiency virus type 1 (HIV-1) nucleoside reverse transcriptase inhibitors, has a 3'-OH and exhibits remarkable potency against wild-type and drug-resistant HIVs. EFdA triphosphate (EFdA-TP) is unique among nucleoside reverse transcriptase inhibitors because it inhibits HIV-1 reverse transcriptase (RT) with multiple mechanisms. (a) EFdA-TP can block RT as a translocation-defective RT inhibitor that dramatically slows DNA synthesis, acting as a de facto immediate chain terminator. Although non-translocated EFdA-MP-terminated primers can be unblocked, they can be efficiently converted back to the EFdA-MP-terminated form. (b) EFdA-TP can function as a delayed chain terminator, allowing incorporation of an additional dNTP before blocking DNA synthesis. In such cases, EFdA-MP-terminated primers are protected from excision. (c) EFdA-MP can be efficiently misincorporated by RT, leading to mismatched primers that are extremely hard to extend and are also protected from excision. The context of template sequence defines the relative contribution of each mechanism and affects the affinity of EFdA-MP for potential incorporation sites, explaining in part the lack of antagonism between EFdA and tenofovir. Changes in the type of nucleotide before EFdA-MP incorporation can alter its mechanism of inhibition from delayed chain terminator to immediate chain terminator. The versatility of EFdA in inhibiting HIV replication by multiple mechanisms may explain why resistance to EFdA is more difficult to emerge.
Keywords: AIDS; Antivirals; EFdA; Enzyme Inhibitor; Human Immunodeficiency Virus (HIV); NRTIs; Nucleoside/Nucleotide Analogue; Reverse Transcriptase; Reverse Transcription.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Hammer S. M., Saag M. S., Schechter M., Montaner J. S., Schooley R. T., Jacobsen D. M., Thompson M. A., Carpenter C. C., Fischl M. A., Gazzard B. G., Gatell J. M., Hirsch M. S., Katzenstein D. A., Richman D. D., Vella S., Yeni P. G., Volberding P. A. (2006) Treatment for adult HIV infection: 2006 recommendations of the International AIDS Society–U.S.A. panel. Top. HIV Med. 14, 827–843 - PubMed
-
- Parniak M. A., Sluis-Cremer N. (2000) Inhibitors of HIV-1 reverse transcriptase. Adv. Pharmacol. 49, 67–109 - PubMed
-
- De Clercq E. (2007) Anti-HIV drugs. Verh. K. Acad. Geneeskd. Belg. 69, 81–104 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01AI076119-S1/AI/NIAID NIH HHS/United States
- R01AI099284/AI/NIAID NIH HHS/United States
- P01GM103368/GM/NIGMS NIH HHS/United States
- R21AI112417/AI/NIAID NIH HHS/United States
- R01 AI100890/AI/NIAID NIH HHS/United States
- R01AI100890/AI/NIAID NIH HHS/United States
- R01AI076119-02S1/AI/NIAID NIH HHS/United States
- P50 GM103368/GM/NIGMS NIH HHS/United States
- R33 AI079801/AI/NIAID NIH HHS/United States
- AI079801/AI/NIAID NIH HHS/United States
- R21 AI079801/AI/NIAID NIH HHS/United States
- R01AI076119/AI/NIAID NIH HHS/United States
- R37 AI076119/AI/NIAID NIH HHS/United States
- R01 AI099284/AI/NIAID NIH HHS/United States
- R21 AI112417/AI/NIAID NIH HHS/United States
- R01 AI076119/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
