

Symmetry breakage in the vertebrate embryo: when does it happen and how does it work?
- PMID: 24972089
- PMCID: PMC4481729
- DOI: 10.1016/j.ydbio.2014.06.014
Symmetry breakage in the vertebrate embryo: when does it happen and how does it work?
Abstract
Asymmetric development of the vertebrate embryo has fascinated embryologists for over a century. Much has been learned since the asymmetric Nodal signaling cascade in the left lateral plate mesoderm was detected, and began to be unraveled over the past decade or two. When and how symmetry is initially broken, however, has remained a matter of debate. Two essentially mutually exclusive models prevail. Cilia-driven leftward flow of extracellular fluids occurs in mammalian, fish and amphibian embryos. A great deal of experimental evidence indicates that this flow is indeed required for symmetry breaking. An alternative model has argued, however, that flow simply acts as an amplification step for early asymmetric cues generated by ion flux during the first cleavage divisions. In this review we critically evaluate the experimental basis of both models. Although a number of open questions persist, the available evidence is best compatible with flow-based symmetry breakage as the archetypical mode of symmetry breakage.
Keywords: Cilia; Ion-flux model; Leftward flow; Left–right asymmetry; Symmetry breakage.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
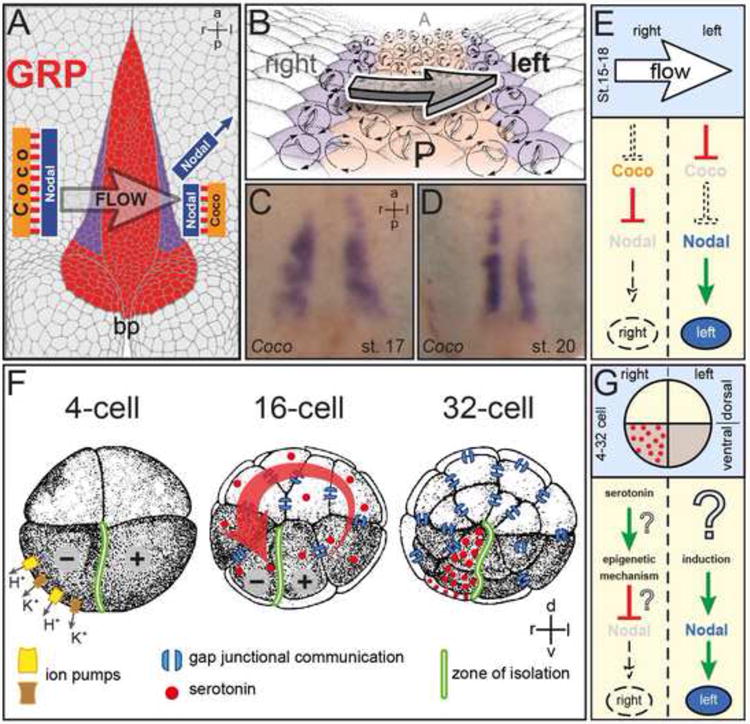
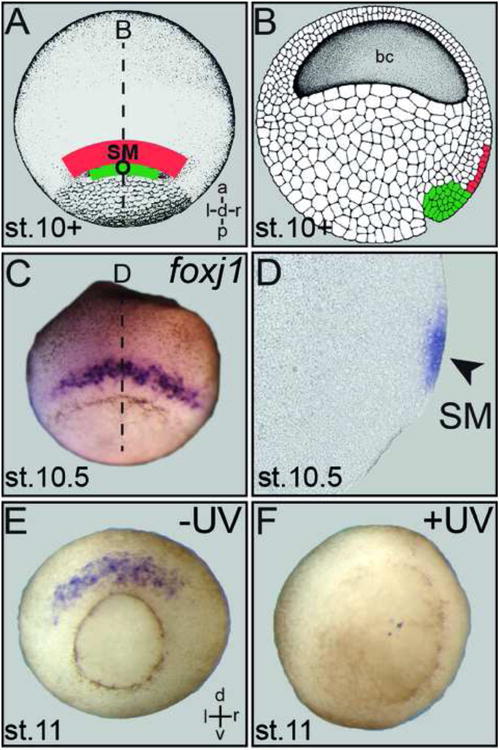
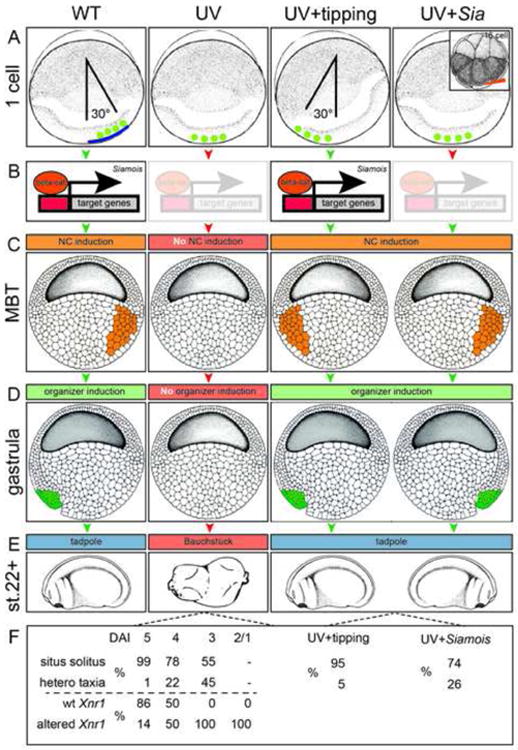
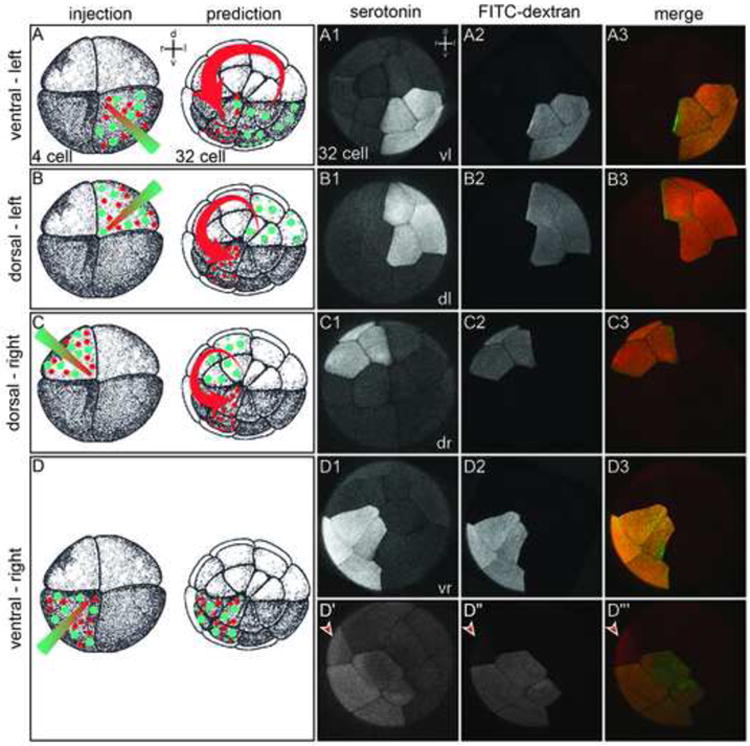
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
