

Increased microtubule assembly rates influence chromosomal instability in colorectal cancer cells
- PMID: 24976383
- PMCID: PMC4389786
- DOI: 10.1038/ncb2994
Increased microtubule assembly rates influence chromosomal instability in colorectal cancer cells
Abstract
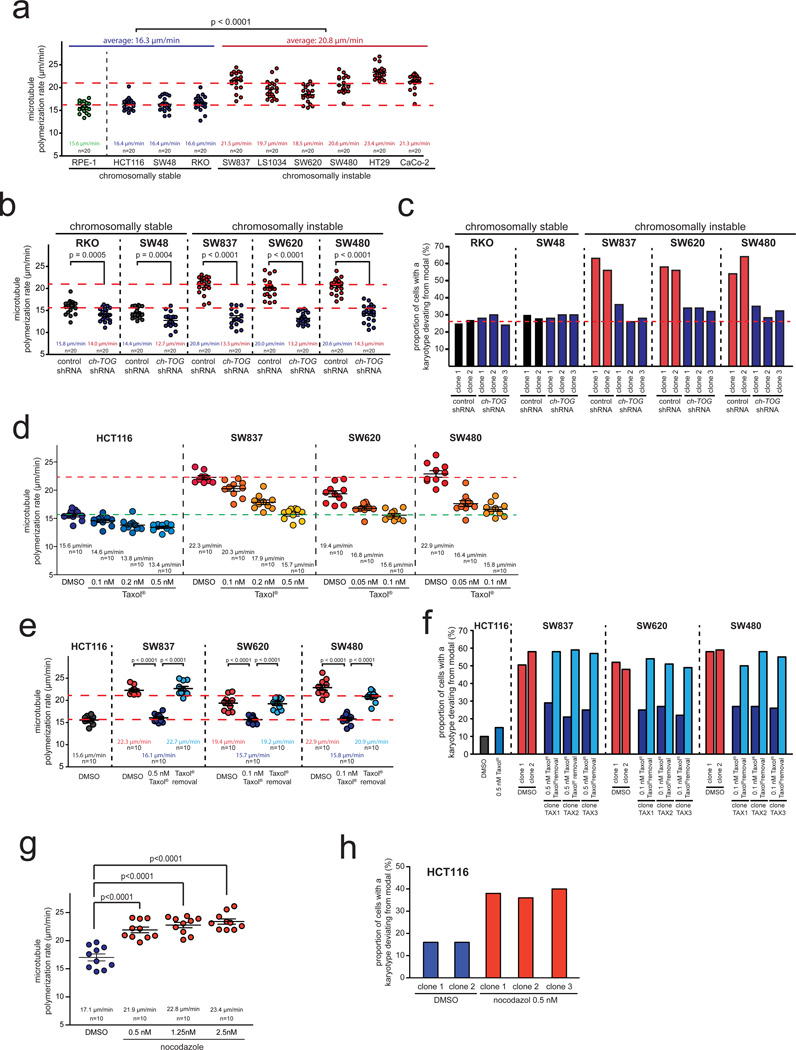
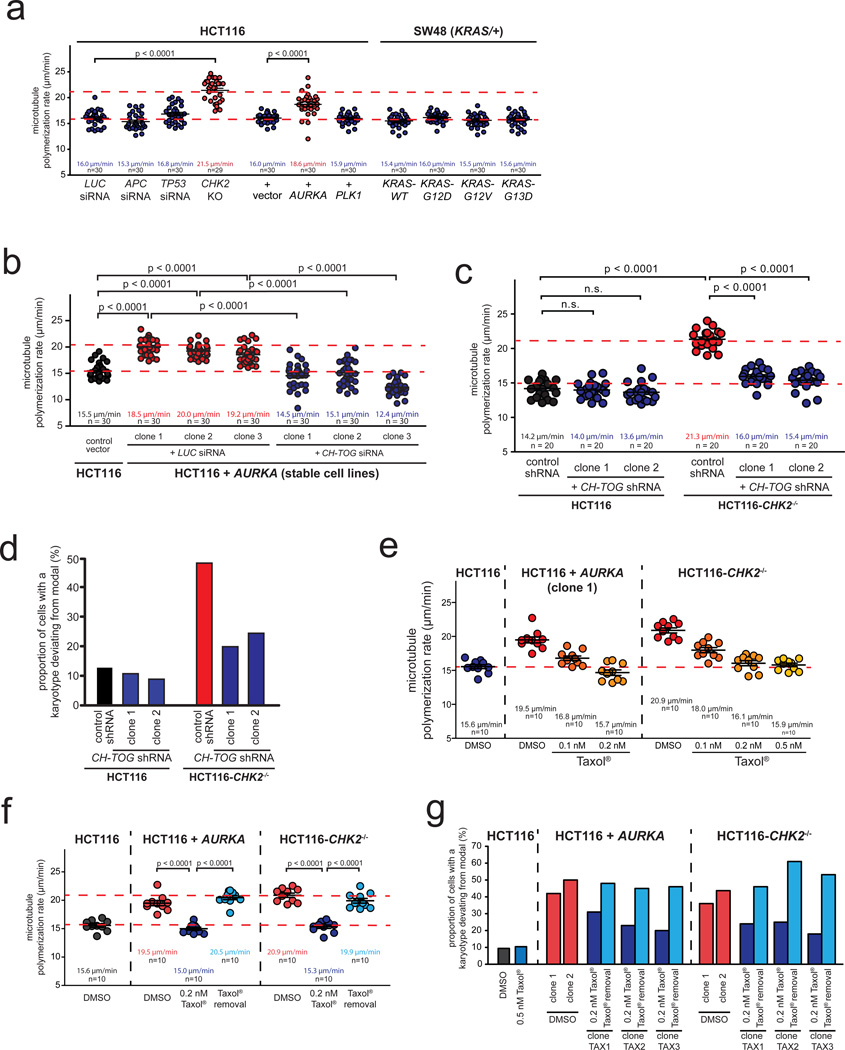
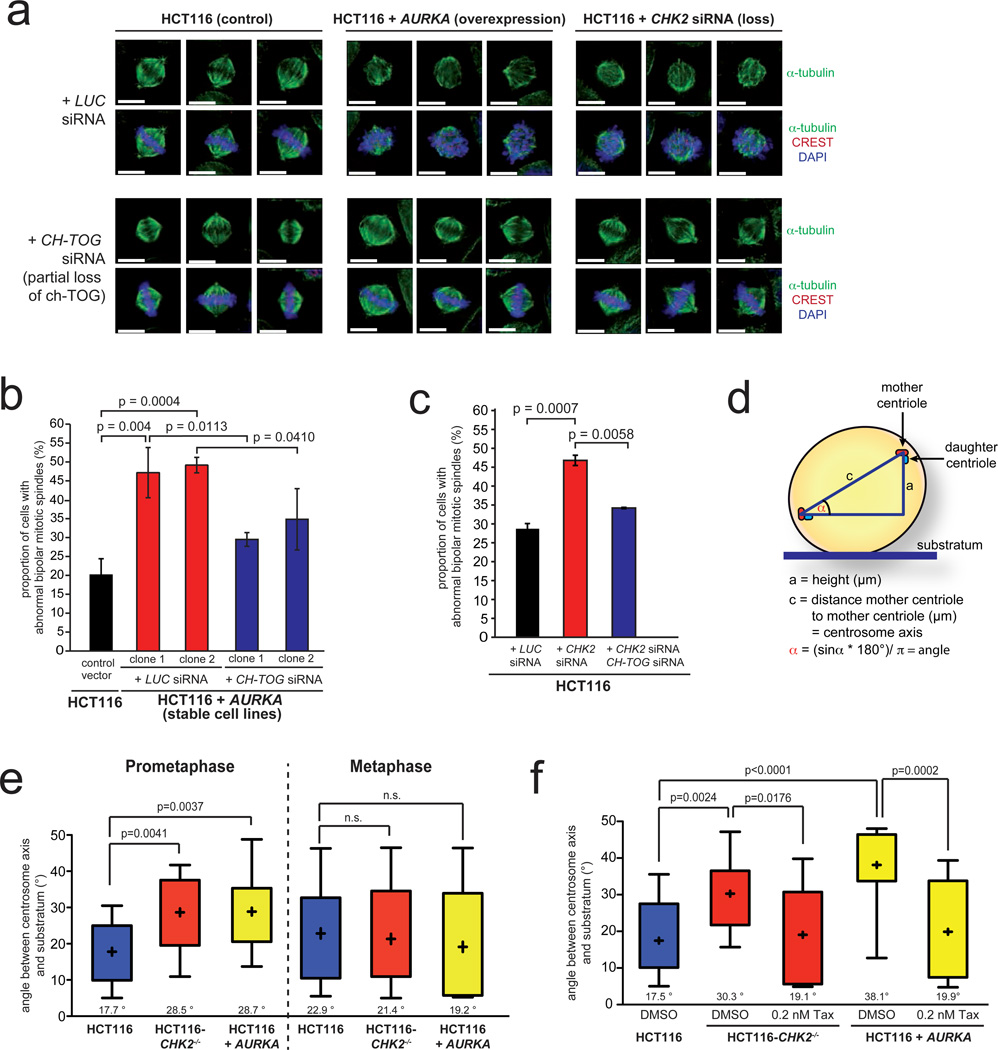
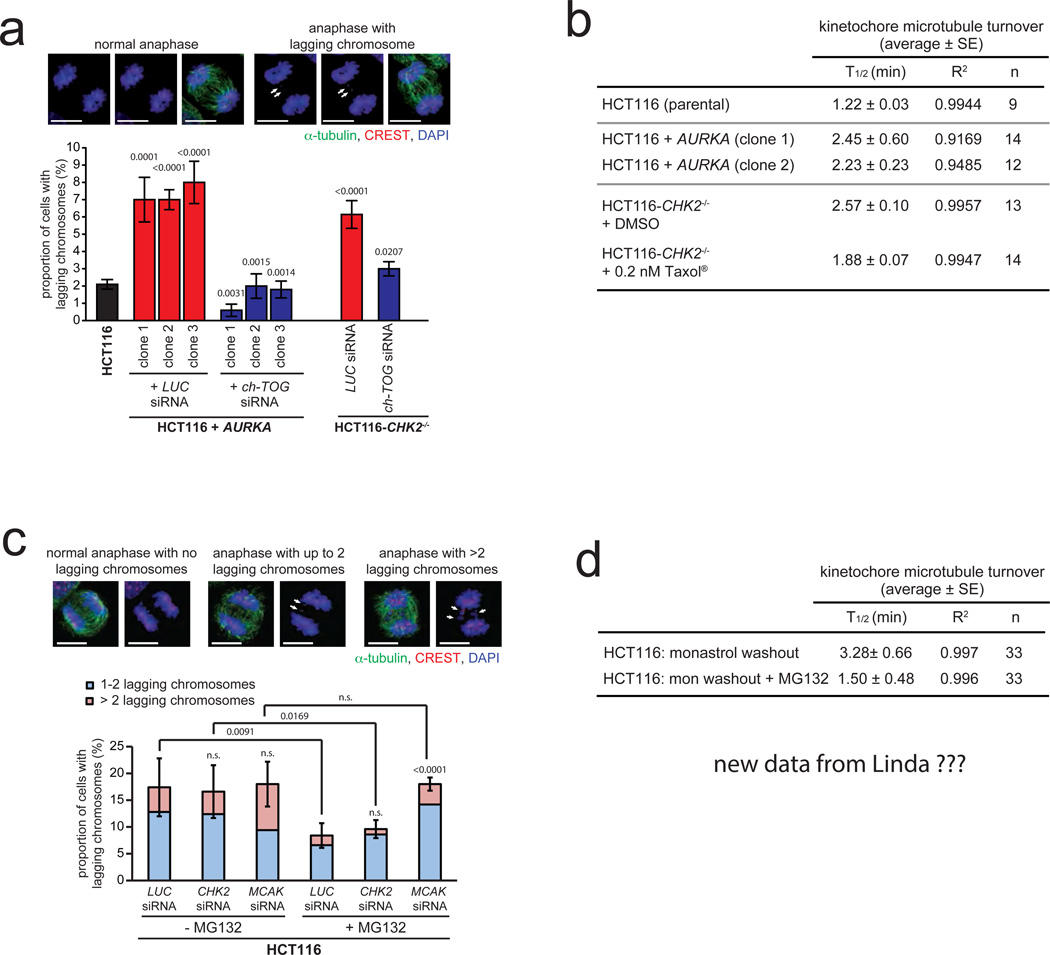
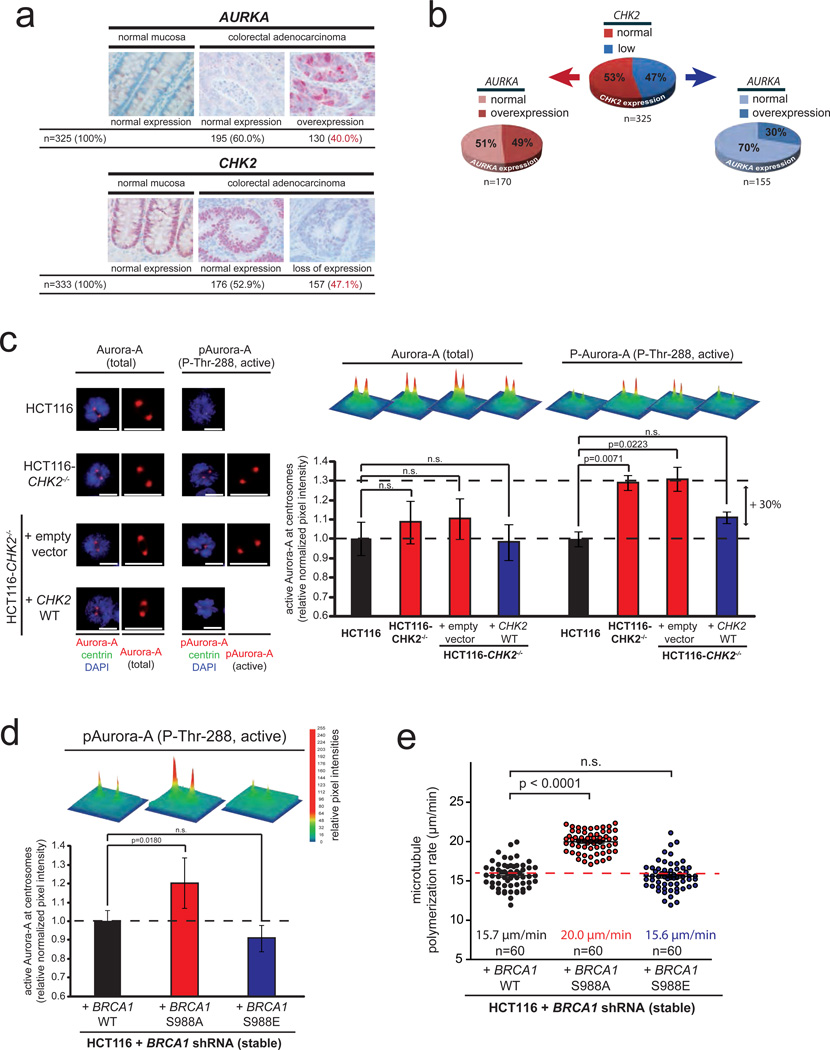
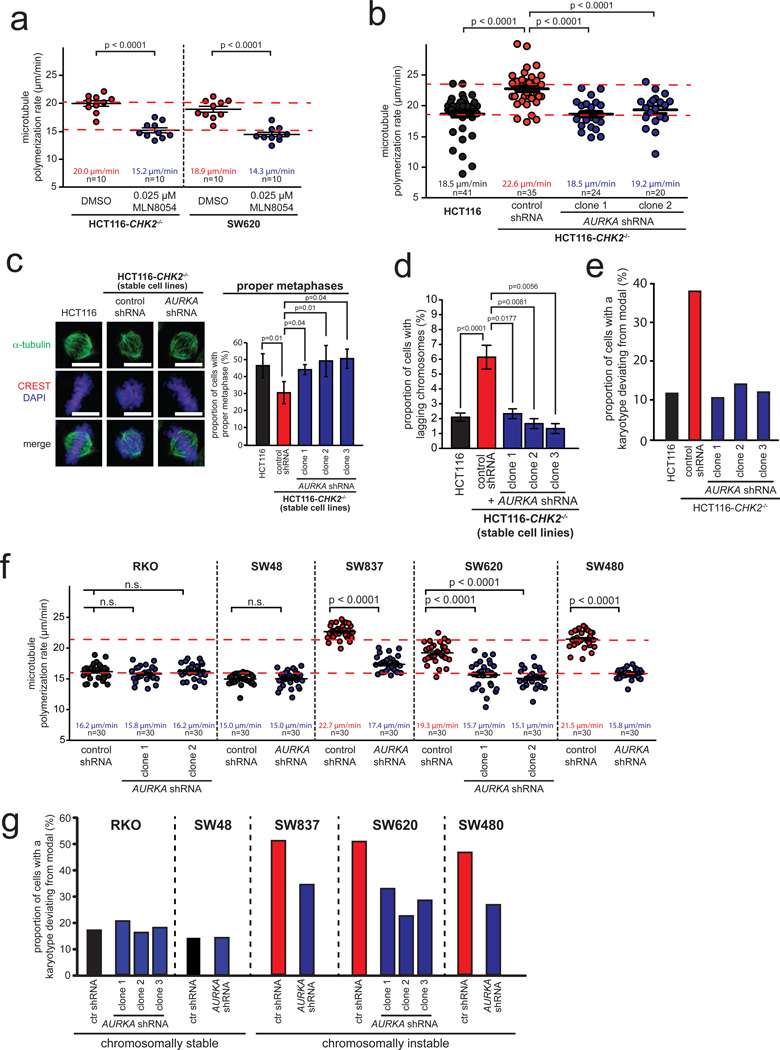
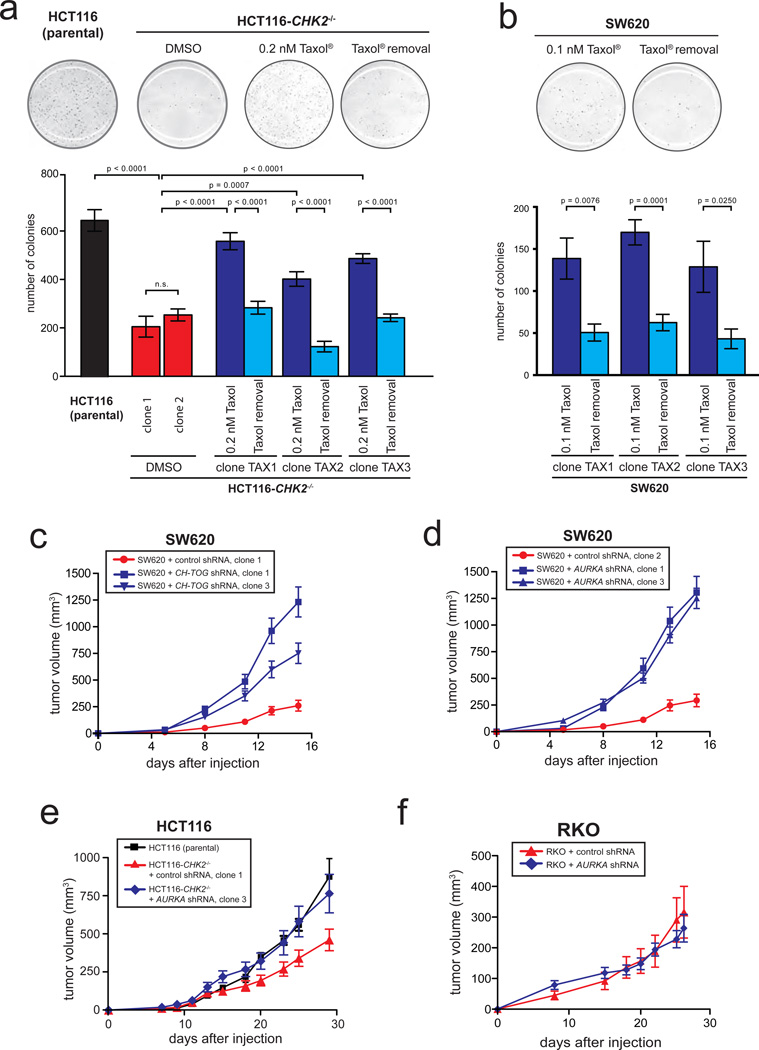
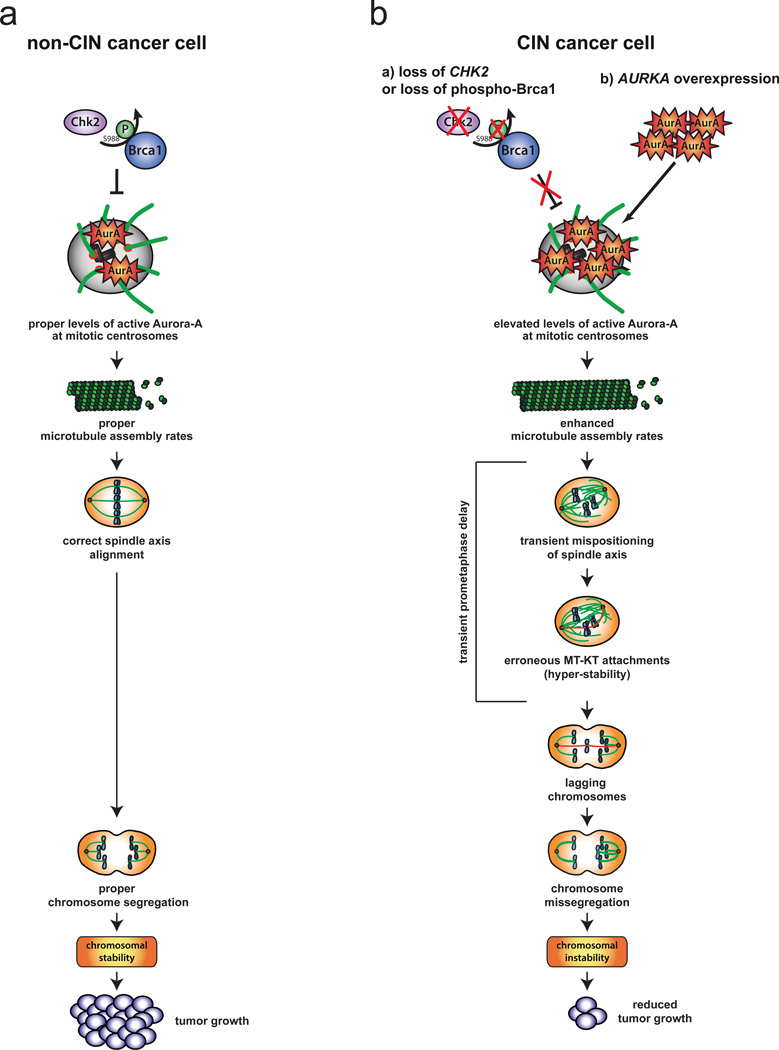
Chromosomal instability (CIN) is defined as the perpetual missegregation of whole chromosomes during mitosis and represents a hallmark of human cancer. However, the mechanisms influencing CIN and its consequences on tumour growth are largely unknown. We identified an increase in microtubule plus-end assembly rates as a mechanism influencing CIN in colorectal cancer cells. This phenotype is induced by overexpression of the oncogene AURKA or by loss of the tumour suppressor gene CHK2, a genetic constitution found in 73% of human colorectal cancers. Increased microtubule assembly rates are associated with transient abnormalities in mitotic spindle geometry promoting the generation of lagging chromosomes and influencing CIN. Reconstitution of proper microtubule assembly rates by chemical or genetic means suppresses CIN and thereby, unexpectedly, accelerates tumour growth in vitro and in vivo. Thus, we identify a fundamental mechanism influencing CIN in cancer cells and reveal its adverse consequence on tumour growth.
Figures
Comment in
-
A growing role for Aurora A in chromosome instability.Nat Cell Biol. 2014 Aug;16(8):739-41. doi: 10.1038/ncb3018. Nat Cell Biol. 2014. PMID: 25082197
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
