

An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore
- PMID: 24979777
- PMCID: PMC4115574
- DOI: 10.1073/pnas.1401591111
An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore
Abstract
Mitochondria maintain tight regulation of inner mitochondrial membrane (IMM) permeability to sustain ATP production. Stressful events cause cellular calcium (Ca(2+)) dysregulation followed by rapid loss of IMM potential known as permeability transition (PT), which produces osmotic shifts, metabolic dysfunction, and cell death. The molecular identity of the mitochondrial PT pore (mPTP) was previously unknown. We show that the purified reconstituted c-subunit ring of the FO of the F1FO ATP synthase forms a voltage-sensitive channel, the persistent opening of which leads to rapid and uncontrolled depolarization of the IMM in cells. Prolonged high matrix Ca(2+) enlarges the c-subunit ring and unhooks it from cyclophilin D/cyclosporine A binding sites in the ATP synthase F1, providing a mechanism for mPTP opening. In contrast, recombinant F1 beta-subunit applied exogenously to the purified c-subunit enhances the probability of pore closure. Depletion of the c-subunit attenuates Ca(2+)-induced IMM depolarization and inhibits Ca(2+) and reactive oxygen species-induced cell death whereas increasing the expression or single-channel conductance of the c-subunit sensitizes to death. We conclude that a highly regulated c-subunit leak channel is a candidate for the mPTP. Beyond cell death, these findings also imply that increasing the probability of c-subunit channel closure in a healthy cell will enhance IMM coupling and increase cellular metabolic efficiency.
Keywords: apoptosis; excitotoxicity; ion channel; metabolism; necrosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

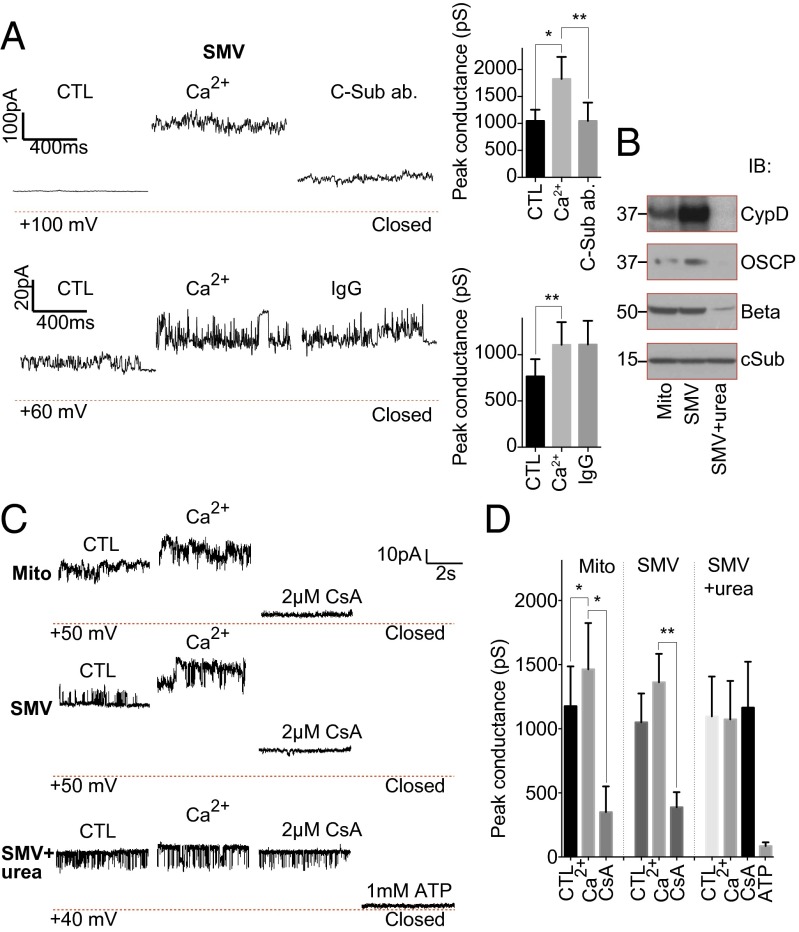

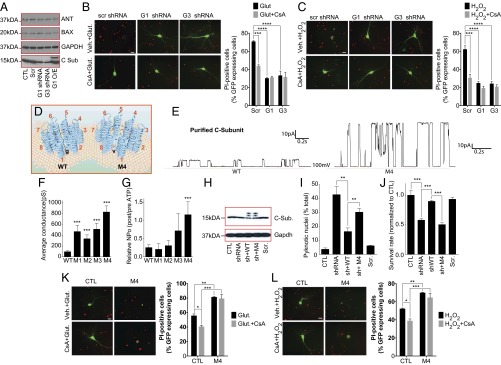
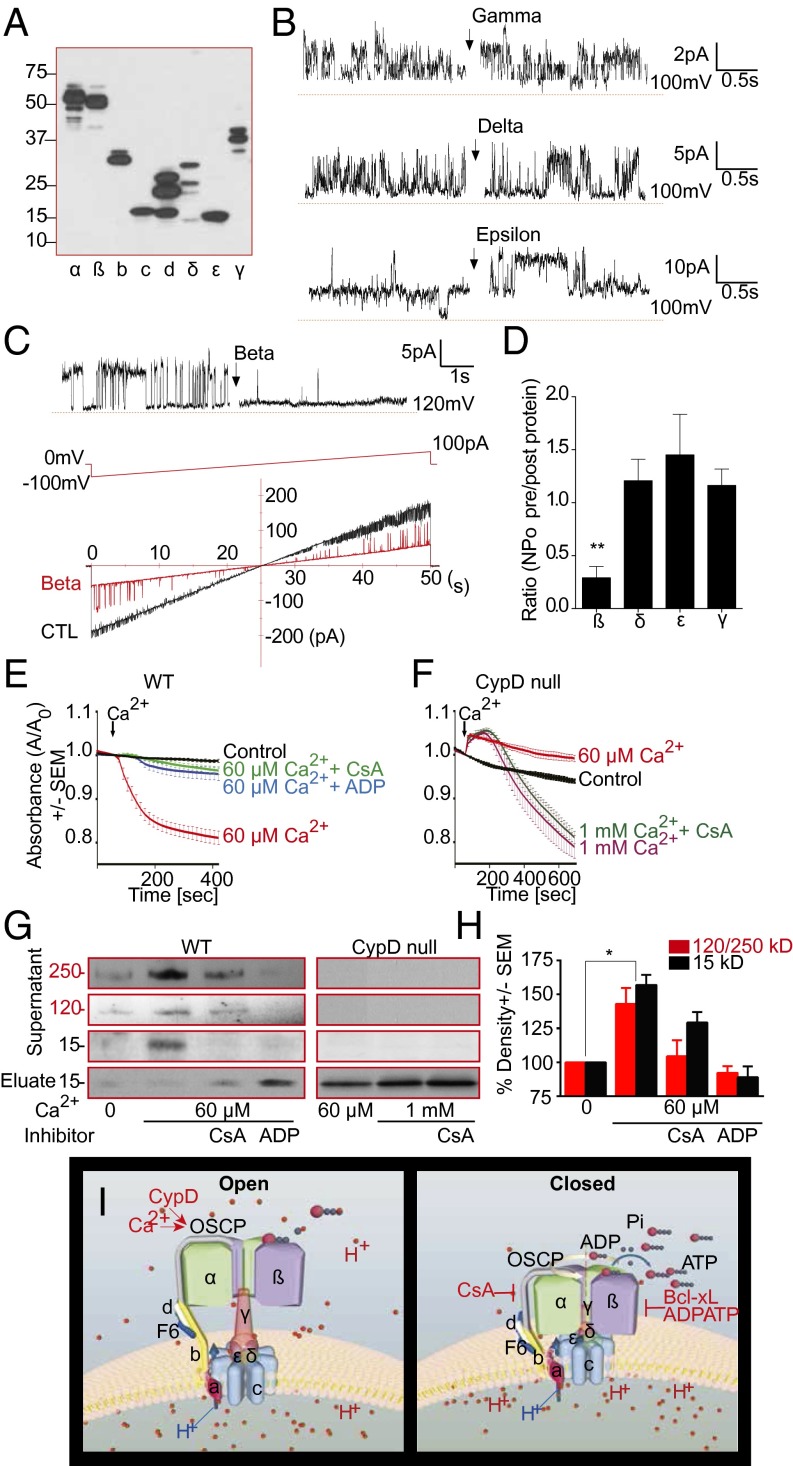
Comment in
-
Identifying the components of the elusive mitochondrial permeability transition pore.Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):10396-7. doi: 10.1073/pnas.1410104111. Epub 2014 Jul 7. Proc Natl Acad Sci U S A. 2014. PMID: 25002521 Free PMC article. No abstract available.
References
-
- Petronilli V, Szabò I, Zoratti M. The inner mitochondrial membrane contains ion-conducting channels similar to those found in bacteria. FEBS Lett. 1989;259(1):137–143. - PubMed
-
- Antonenko YN, Smith D, Kinnally KW, Tedeschi H. Single-channel activity induced in mitoplasts by alkaline pH. Biochim Biophys Acta. 1994;1194(2):247–254. - PubMed
-
- Bernardi P, et al. Modulation of the mitochondrial permeability transition pore: Effect of protons and divalent cations. J Biol Chem. 1992;267(5):2934–2939. - PubMed
-
- Bernardi P. Modulation of the mitochondrial cyclosporin A-sensitive permeability transition pore by the proton electrochemical gradient: Evidence that the pore can be opened by membrane depolarization. J Biol Chem. 1992;267(13):8834–8839. - PubMed
-
- Jonas EA, Buchanan J, Kaczmarek LK. Prolonged activation of mitochondrial conductances during synaptic transmission. Science. 1999;286(5443):1347–1350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
