

Structure of the C. elegans ZYG-1 cryptic polo box suggests a conserved mechanism for centriolar docking of Plk4 kinases
- PMID: 24980795
- PMCID: PMC4126857
- DOI: 10.1016/j.str.2014.05.009
Structure of the C. elegans ZYG-1 cryptic polo box suggests a conserved mechanism for centriolar docking of Plk4 kinases
Abstract
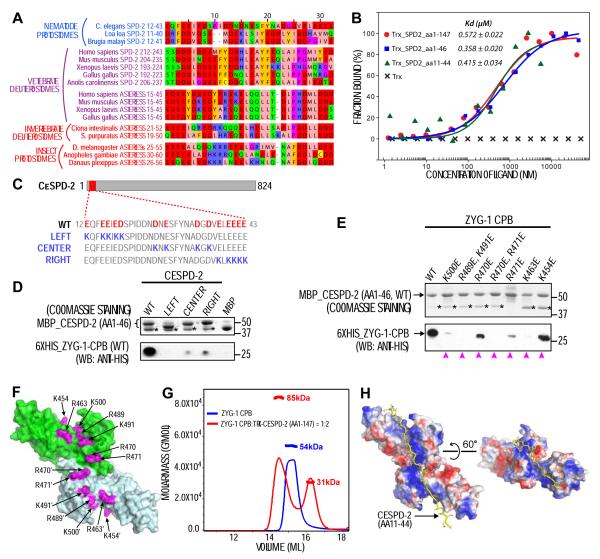
Plk4 family kinases control centriole assembly. Plk4s target mother centrioles through an interaction between their cryptic polo box (CPB) and acidic regions in the centriolar receptors SPD-2/Cep192 and/or Asterless/Cep152. Here, we report a crystal structure for the CPB of C. elegans ZYG-1, which forms a Z-shaped dimer containing an intermolecular β sheet with an extended basic surface patch. Biochemical and in vivo analysis revealed that electrostatic interactions dock the ZYG-1 CPB basic patch onto the SPD-2-derived acidic region to promote ZYG-1 targeting and new centriole assembly. Analysis of a different crystal form of the Drosophila Plk4 (DmPlk4) CPB suggests that it also forms a Z-shaped dimer. Comparison of the ZYG-1 and DmPlk4 CPBs revealed structural changes in the ZYG-1 CPB that confer selectivity for binding SPD-2 over Asterless-derived acidic regions. Overall, our findings suggest a conserved mechanism for centriolar docking of Plk4 homologs that initiate daughter centriole assembly.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures






Comment in
-
Polo-like kinase 4 shapes up.Structure. 2014 Aug 5;22(8):1071-1073. doi: 10.1016/j.str.2014.07.004. Structure. 2014. PMID: 25099950
References
-
- Anderhub SJ, Krämer A, Maier B. Centrosome amplification in tumorigenesis. Cancer Lett. 2012;322:8–17. doi:10.1016/j.canlet.2012.02.006. - PubMed
-
- Brito DA, Gouveia SM, Bettencourt-Dias M. Deconstructing the centriole: structure and number control. Curr. Opin. Cell Biol. 2012;24:4–13. doi:10.1016/j.ceb.2012.01.003. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
